Explore
Explore Validate
Validate Learn
Learn44-768G
antibody from Invitrogen Antibodies
Targeting: MAPT
DDPAC, FLJ31424, FTDP-17, MAPTL, MGC138549, MSTD, MTBT1, MTBT2, PPND, PPP1R103, tau
 Western blot
Western blotAntibody data
- Antibody Data
- Antigen structure
- References [25]
- Comments [0]
- Validations
- Western blot [1]
- Immunocytochemistry [1]
- Immunohistochemistry [1]
- Flow cytometry [1]
- Other assay [9]
Submit
Validation data
Reference
Comment
Report error
- Product number
- 44-768G - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- Phospho-Tau (Ser199, Ser202) Polyclonal Antibody
- Antibody type
- Polyclonal
- Antigen
- Synthetic peptide
- Description
- This antibody has been negatively preadsorbed using a non-phosphopeptide corresponding to the site of phosphorylation to remove antibody that is reactive with non-phosphorylated tau. The final product is generated by affinity chromatography using a tau-derived peptide that is phosphorylated at serines 199 and 202.
Submitted references High-fat diet exacerbates cognitive decline in mouse models of Alzheimer's disease and mixed dementia in a sex-dependent manner.
Tau phosphorylation and OPA1 proteolysis are unrelated events: Implications for Alzheimer's Disease.
PKR kinase directly regulates tau expression and Alzheimer's disease-related tau phosphorylation.
Phosphorylated tau interactome in the human Alzheimer's disease brain.
Noncanonical function of an autophagy protein prevents spontaneous Alzheimer's disease.
DNA repair deficiency and senescence in concussed professional athletes involved in contact sports.
DNA damage as a marker of brain damage in individuals with history of concussions.
LC3-Associated Endocytosis Facilitates β-Amyloid Clearance and Mitigates Neurodegeneration in Murine Alzheimer's Disease.
A post-translational modification signature defines changes in soluble tau correlating with oligomerization in early stage Alzheimer's disease brain.
Tau secretion is correlated to an increase of Golgi dynamics.
A validated antibody panel for the characterization of tau post-translational modifications.
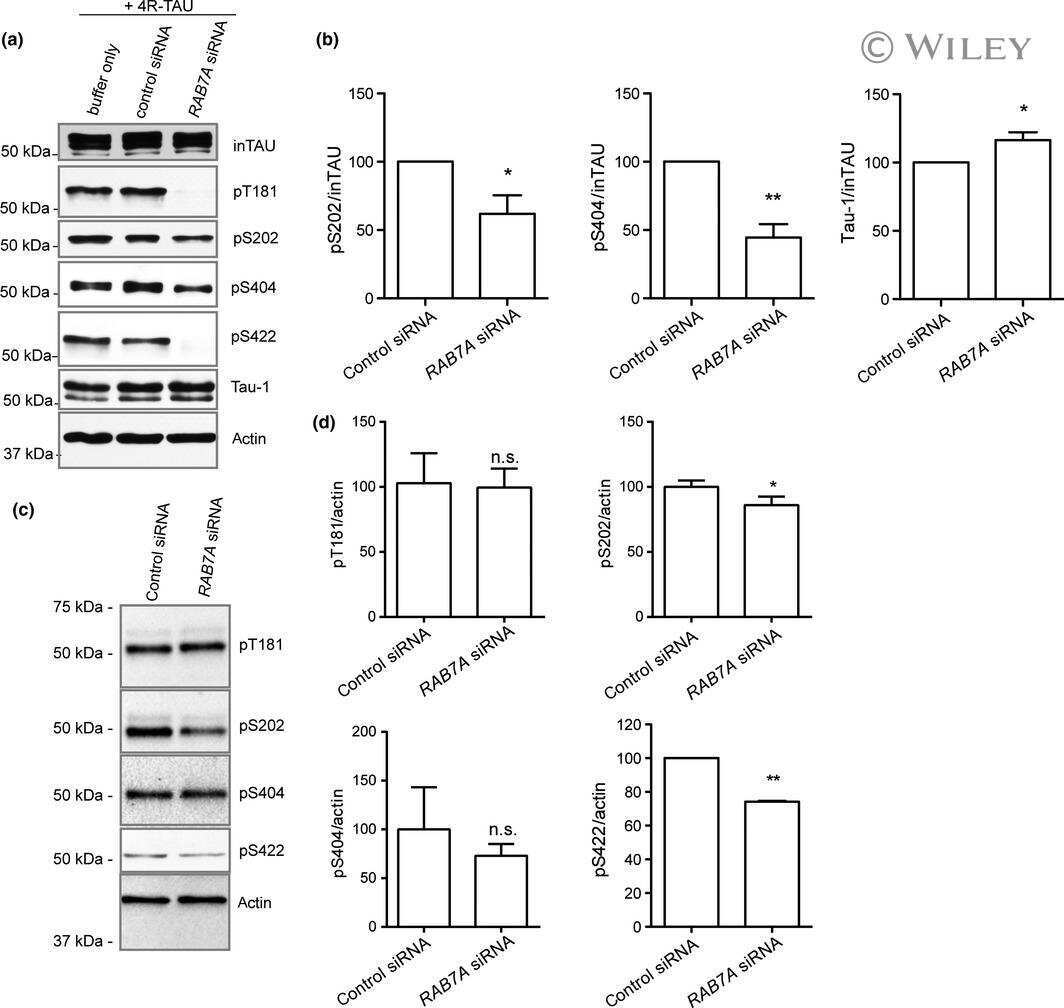
Rab7A regulates tau secretion.
Liraglutide Improves Water Maze Learning and Memory Performance While Reduces Hyperphosphorylation of Tau and Neurofilaments in APP/PS1/Tau Triple Transgenic Mice.
Chronic Traumatic Encephalopathy-Like Abnormalities in a Routine Neuropathology Service.
The choice of general anesthetics may not affect neuroinflammation and impairment of learning and memory after surgery in elderly rats.
Fyn inhibition rescues established memory and synapse loss in Alzheimer mice.
Human APOE genotype affects intraneuronal Aβ1-42 accumulation in a lentiviral gene transfer model.
Cellular prion protein modulates β-amyloid deposition in aged APP/PS1 transgenic mice.
Transient focal cerebral ischemia/reperfusion induces early and chronic axonal changes in rats: its importance for the risk of Alzheimer's disease.
Lidocaine attenuates cognitive impairment after isoflurane anesthesia in old rats.
Limiting multiple sclerosis related axonopathy by blocking Nogo receptor and CRMP-2 phosphorylation.
Use of inhibitors in the study of MAP kinases.
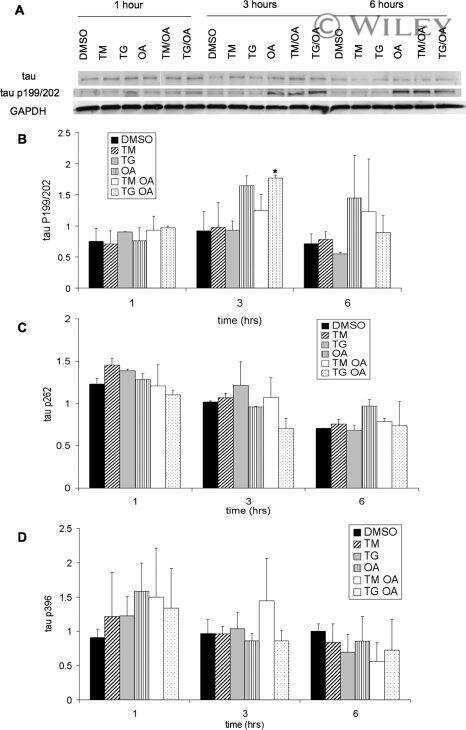
Transgenic mouse and cell culture models demonstrate a lack of mechanistic connection between endoplasmic reticulum stress and tau dysfunction.
Increased tau phosphorylation and cleavage in mouse models of type 1 and type 2 diabetes.
A Cdk5 inhibitory peptide reduces tau hyperphosphorylation and apoptosis in neurons.
Gannon OJ, Robison LS, Salinero AE, Abi-Ghanem C, Mansour FM, Kelly RD, Tyagi A, Brawley RR, Ogg JD, Zuloaga KL
Journal of neuroinflammation 2022 May 14;19(1):110
Journal of neuroinflammation 2022 May 14;19(1):110
Tau phosphorylation and OPA1 proteolysis are unrelated events: Implications for Alzheimer's Disease.
Alavi MV
Biochimica et biophysica acta. Molecular cell research 2021 Nov;1868(12):119116
Biochimica et biophysica acta. Molecular cell research 2021 Nov;1868(12):119116
PKR kinase directly regulates tau expression and Alzheimer's disease-related tau phosphorylation.
Reimer L, Betzer C, Kofoed RH, Volbracht C, Fog K, Kurhade C, Nilsson E, Överby AK, Jensen PH
Brain pathology (Zurich, Switzerland) 2021 Jan;31(1):103-119
Brain pathology (Zurich, Switzerland) 2021 Jan;31(1):103-119
Phosphorylated tau interactome in the human Alzheimer's disease brain.
Drummond E, Pires G, MacMurray C, Askenazi M, Nayak S, Bourdon M, Safar J, Ueberheide B, Wisniewski T
Brain : a journal of neurology 2020 Sep 1;143(9):2803-2817
Brain : a journal of neurology 2020 Sep 1;143(9):2803-2817
Noncanonical function of an autophagy protein prevents spontaneous Alzheimer's disease.
Heckmann BL, Teubner BJW, Boada-Romero E, Tummers B, Guy C, Fitzgerald P, Mayer U, Carding S, Zakharenko SS, Wileman T, Green DR
Science advances 2020 Aug;6(33):eabb9036
Science advances 2020 Aug;6(33):eabb9036
DNA repair deficiency and senescence in concussed professional athletes involved in contact sports.
Schwab N, Grenier K, Hazrati LN
Acta neuropathologica communications 2019 Nov 14;7(1):182
Acta neuropathologica communications 2019 Nov 14;7(1):182
DNA damage as a marker of brain damage in individuals with history of concussions.
Schwab N, Tator C, Hazrati LN
Laboratory investigation; a journal of technical methods and pathology 2019 Jul;99(7):1008-1018
Laboratory investigation; a journal of technical methods and pathology 2019 Jul;99(7):1008-1018
LC3-Associated Endocytosis Facilitates β-Amyloid Clearance and Mitigates Neurodegeneration in Murine Alzheimer's Disease.
Heckmann BL, Teubner BJW, Tummers B, Boada-Romero E, Harris L, Yang M, Guy CS, Zakharenko SS, Green DR
Cell 2019 Jul 25;178(3):536-551.e14
Cell 2019 Jul 25;178(3):536-551.e14
A post-translational modification signature defines changes in soluble tau correlating with oligomerization in early stage Alzheimer's disease brain.
Ercan-Herbst E, Ehrig J, Schöndorf DC, Behrendt A, Klaus B, Gomez Ramos B, Prat Oriol N, Weber C, Ehrnhoefer DE
Acta neuropathologica communications 2019 Dec 3;7(1):192
Acta neuropathologica communications 2019 Dec 3;7(1):192
Tau secretion is correlated to an increase of Golgi dynamics.
Mohamed NV, Desjardins A, Leclerc N
PloS one 2017;12(5):e0178288
PloS one 2017;12(5):e0178288
A validated antibody panel for the characterization of tau post-translational modifications.
Ercan E, Eid S, Weber C, Kowalski A, Bichmann M, Behrendt A, Matthes F, Krauss S, Reinhardt P, Fulle S, Ehrnhoefer DE
Molecular neurodegeneration 2017 Nov 21;12(1):87
Molecular neurodegeneration 2017 Nov 21;12(1):87
Rab7A regulates tau secretion.
Rodriguez L, Mohamed NV, Desjardins A, Lippé R, Fon EA, Leclerc N
Journal of neurochemistry 2017 May;141(4):592-605
Journal of neurochemistry 2017 May;141(4):592-605
Liraglutide Improves Water Maze Learning and Memory Performance While Reduces Hyperphosphorylation of Tau and Neurofilaments in APP/PS1/Tau Triple Transgenic Mice.
Chen S, Sun J, Zhao G, Guo A, Chen Y, Fu R, Deng Y
Neurochemical research 2017 Aug;42(8):2326-2335
Neurochemical research 2017 Aug;42(8):2326-2335
Chronic Traumatic Encephalopathy-Like Abnormalities in a Routine Neuropathology Service.
Noy S, Krawitz S, Del Bigio MR
Journal of neuropathology and experimental neurology 2016 Dec 1;75(12):1145-1154
Journal of neuropathology and experimental neurology 2016 Dec 1;75(12):1145-1154
The choice of general anesthetics may not affect neuroinflammation and impairment of learning and memory after surgery in elderly rats.
Zhang J, Tan H, Jiang W, Zuo Z
Journal of neuroimmune pharmacology : the official journal of the Society on NeuroImmune Pharmacology 2015 Mar;10(1):179-89
Journal of neuroimmune pharmacology : the official journal of the Society on NeuroImmune Pharmacology 2015 Mar;10(1):179-89
Fyn inhibition rescues established memory and synapse loss in Alzheimer mice.
Kaufman AC, Salazar SV, Haas LT, Yang J, Kostylev MA, Jeng AT, Robinson SA, Gunther EC, van Dyck CH, Nygaard HB, Strittmatter SM
Annals of neurology 2015 Jun;77(6):953-71
Annals of neurology 2015 Jun;77(6):953-71
Human APOE genotype affects intraneuronal Aβ1-42 accumulation in a lentiviral gene transfer model.
Zhao W, Dumanis SB, Tamboli IY, Rodriguez GA, Jo Ladu M, Moussa CE, William Rebeck G
Human molecular genetics 2014 Mar 1;23(5):1365-75
Human molecular genetics 2014 Mar 1;23(5):1365-75
Cellular prion protein modulates β-amyloid deposition in aged APP/PS1 transgenic mice.
Ordóñez-Gutiérrez L, Torres JM, Gavín R, Antón M, Arroba-Espinosa AI, Espinosa JC, Vergara C, Del Río JA, Wandosell F
Neurobiology of aging 2013 Dec;34(12):2793-804
Neurobiology of aging 2013 Dec;34(12):2793-804
Transient focal cerebral ischemia/reperfusion induces early and chronic axonal changes in rats: its importance for the risk of Alzheimer's disease.
Zhang Q, Gao T, Luo Y, Chen X, Gao G, Gao X, Zhou Y, Dai J
PloS one 2012;7(3):e33722
PloS one 2012;7(3):e33722
Lidocaine attenuates cognitive impairment after isoflurane anesthesia in old rats.
Lin D, Cao L, Wang Z, Li J, Washington JM, Zuo Z
Behavioural brain research 2012 Mar 17;228(2):319-27
Behavioural brain research 2012 Mar 17;228(2):319-27
Limiting multiple sclerosis related axonopathy by blocking Nogo receptor and CRMP-2 phosphorylation.
Petratos S, Ozturk E, Azari MF, Kenny R, Lee JY, Magee KA, Harvey AR, McDonald C, Taghian K, Moussa L, Mun Aui P, Siatskas C, Litwak S, Fehlings MG, Strittmatter SM, Bernard CC
Brain : a journal of neurology 2012 Jun;135(Pt 6):1794-818
Brain : a journal of neurology 2012 Jun;135(Pt 6):1794-818
Use of inhibitors in the study of MAP kinases.
Burkhard K, Shapiro P
Methods in molecular biology (Clifton, N.J.) 2010;661:107-22
Methods in molecular biology (Clifton, N.J.) 2010;661:107-22
Transgenic mouse and cell culture models demonstrate a lack of mechanistic connection between endoplasmic reticulum stress and tau dysfunction.
Spatara ML, Robinson AS
Journal of neuroscience research 2010 Jul;88(9):1951-61
Journal of neuroscience research 2010 Jul;88(9):1951-61
Increased tau phosphorylation and cleavage in mouse models of type 1 and type 2 diabetes.
Kim B, Backus C, Oh S, Hayes JM, Feldman EL
Endocrinology 2009 Dec;150(12):5294-301
Endocrinology 2009 Dec;150(12):5294-301
A Cdk5 inhibitory peptide reduces tau hyperphosphorylation and apoptosis in neurons.
Zheng YL, Kesavapany S, Gravell M, Hamilton RS, Schubert M, Amin N, Albers W, Grant P, Pant HC
The EMBO journal 2005 Jan 12;24(1):209-20
The EMBO journal 2005 Jan 12;24(1):209-20
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
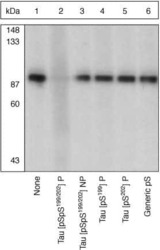
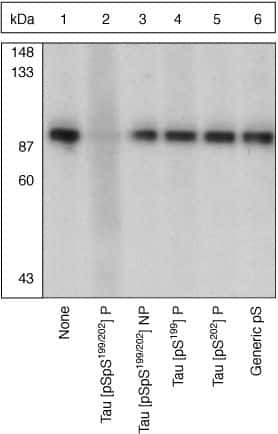
- Experimental details
- Up-regulation and Antibody-Peptide Competition. Peptide Competition. Extracts of from African green monkey kidney (CV-1) cells were resolved by SDS-PAGE on a 10% polyacrylamide gel and transferred to nitrocellulose. Membranes were blocked with a 5% BSA-TBST buffer overnight at 4°C, then incubated with Tau [pSpS199/202] antibody for two hours at room temperature in a 3% BSA-TBST buffer, following prior incubation with: no peptide (1), the phosphopeptide immunogen (2), the non-phosphorylated peptide corresponding to the immunogen (3), the tau phosphopeptide corresponding to [pS199] (4), the tau phosphopeptide corresponding to [pS202] (5), or, a generic phosphoserine-containing peptide (6). After washing, membranes were incubated with goat F(ab’)2 anti-rabbit IgG alkaline phosphatase conjugate (Product # ALI4405) and bands were detected using the Tropix WesternStar™ method.The data show that only the phosphopeptide corresponding to Tau [pSpS199/202] completely blocks the antibody signal, demonstrating the specificity of the antibody. (Cat. No. 44768G).
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
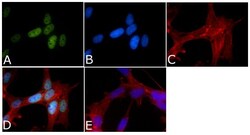
- Experimental details
- Immunofluorescent analysis of Phospho-Tau pSer199/pSer202 Antibody was done on 70% confluent log phase SHSY5Y cells. The cells were fixed with 4% paraformaldehyde for 15 minutes, permeabilized with 0.25% Triton™ X-100 for 10 minutes, and blocked with 5% BSA for 1 hour at room temperature. The cells were labeled with Phospho-Tau pSer199/pSer202 Antibody (Product # 44-768G) at 1µg/mL in 1% BSA and incubated for 3 hours at room temperature and then labeled with Alexa Fluor 488 Goat Anti-Rabbit IgG Secondary Antibody (Product # A-11008) at a dilution of 1:400 for 45 minutes at room temperature (Panel a: green). Nuclei (Panel b: blue) were stained with SlowFade® Gold Antifade Mountant with DAPI (Product # S36938). F-actin (Panel c: red) was stained with Alexa Fluor 594 Phalloidin (Product # A12381). Panel d is a merged image showing nuclear localization. Panel e is a no primary antibody control. The images were captured at 40X magnification.
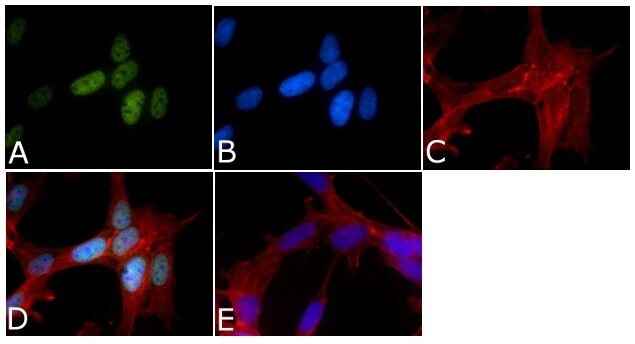
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
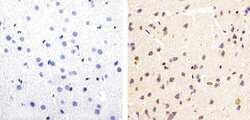
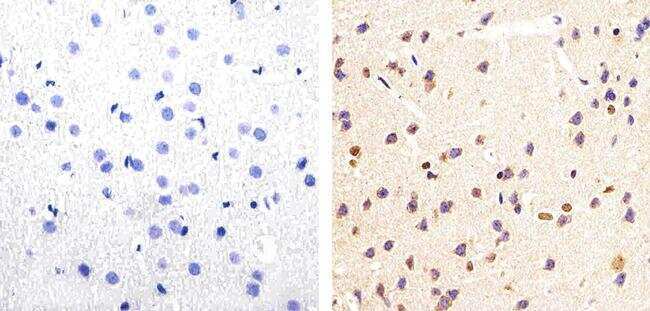
- Experimental details
- Immunohistochemistry analysis of Phospho-Tau (pSpS199/202) showing staining in the cytoplasm of paraffin-embedded mouse brain tissue (right) compared to a negative control without primary antibody (left). To expose target proteins, antigen retrieval was performed using 10mM sodium citrate (pH 6.0), microwaved for 8-15 min. Following antigen retrieval, tissues were blocked in 3% H2O2-methanol for 15 min at room temperature, washed with ddH2O and PBS, and then probed with a Phospho-Tau (pSpS199/202) polyclonal antibody (Product # 44-768G) diluted in 3% BSA-PBS at a dilution of 1:20 overnight at 4ºC in a humidified chamber. Tissues were washed extensively in PBST and detection was performed using an HRP-conjugated secondary antibody followed by colorimetric detection using a DAB kit. Tissues were counterstained with hematoxylin and dehydrated with ethanol and xylene to prep for mounting.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
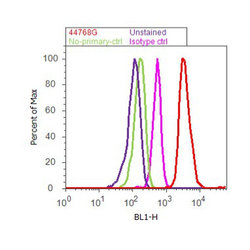
- Experimental details
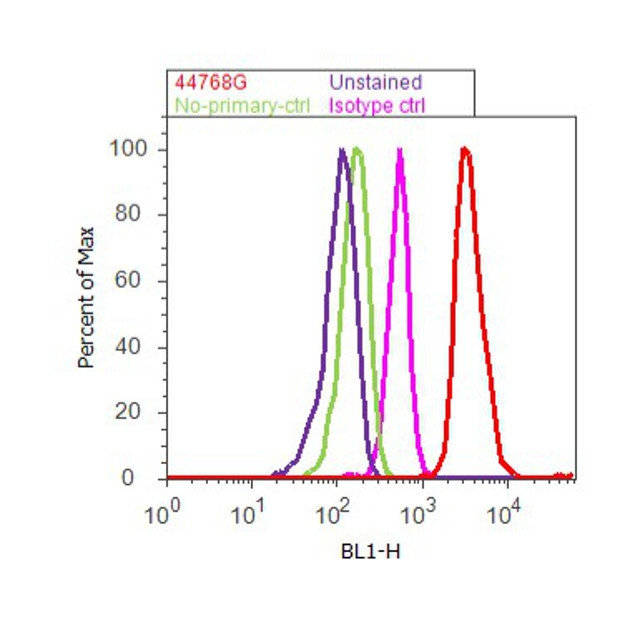
- Flow cytometry analysis of Tau [pSpS199/202] was done on SH-SY5Y cells. Cells were fixed with 70% ethanol for 10 minutes, permeabilized with 0.25% Triton™ X-100 for 20 minutes, and blocked with 5% BSA for 30 minutes at room temperature. Cells were labeled with Tau [pSpS199/202] Rabbit Polyclonal Antibody (44768G, red histogram) or with rabbit isotype control (pink histogram) at 3-5 ug/million cells in 2.5% BSA. After incubation at room temperature for 2 hours, the cells were labeled with Alexa Fluor® 488 Goat Anti-Rabbit Secondary Antibody (A11008) at a dilution of 1:400 for 30 minutes at room temperature. The representative 10,000 cells were acquired and analyzed for each sample using an Attune® Acoustic Focusing Cytometer. The purple histogram represents unstained control cells and the green histogram represents no-primary-antibody control.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
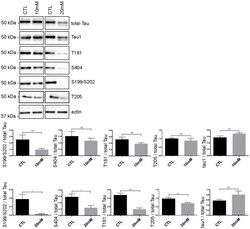
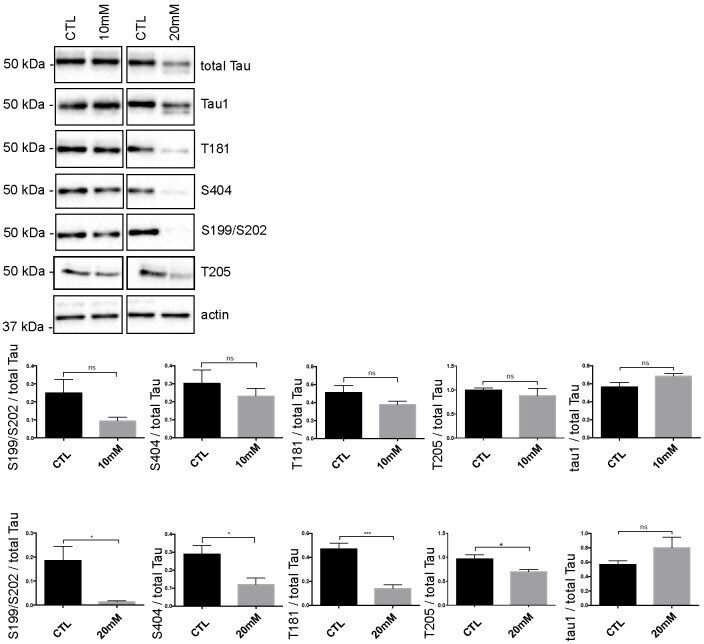
- Fig 5 Tau phosphorylation is decreased in neurons treated with 20 mM KCl. Immunoblot analysis of Tau phosphorylation in neurons treated either with 10mM or 20mM KCl. The phosphorylation of S199/S202, T181, T205 and S404 was not significantly affected by 10mM of KCl treatment whereas the phosphorylation of all these sites was decreased by 20mM of KCl treatment. The signal of the phospho-antibodies was normalized to that of total Tau (n = 5, mean +- SEM, unpaired t-test two-tailed, * P < 0.05, *** P < 0.001). The amount of dephosphorylated Tau revealed by the Tau-1 antibody was normalized to that of total Tau lysate (n = 5, mean +- SEM, unpaired t-test two-tailed, * P < 0.05).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
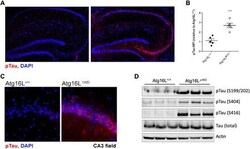
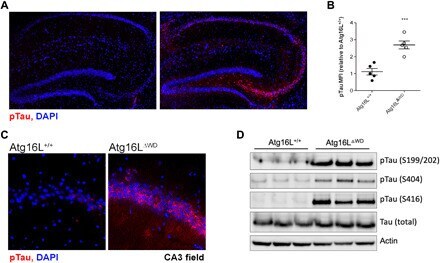
- Fig. 2 Aged mice lacking the WD domain of Atg16L have robust Tau hyperphosphorylation. ( A ) Representative imaging of S199/202 Tau phosphorylation in hippocampus. ( B ) Quantification of Tau phosphorylation in the hippocampus; each point represents and individual mouse. ( C ) CA3 field-specific imaging of S199/202 Tau phosphorylation. ( D ) Immunoblot analysis of whole brain for multiple species of phospho-Tau (pTau) compared to total-Tau. Actin was used as a loading control. *** P < 0.001.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
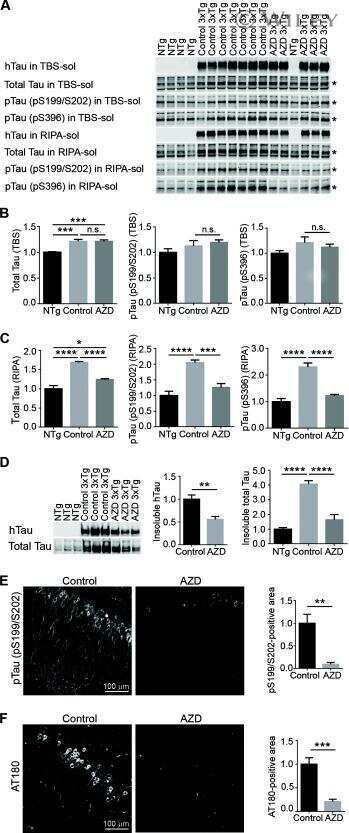
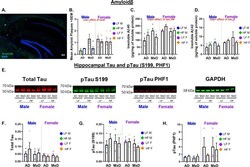
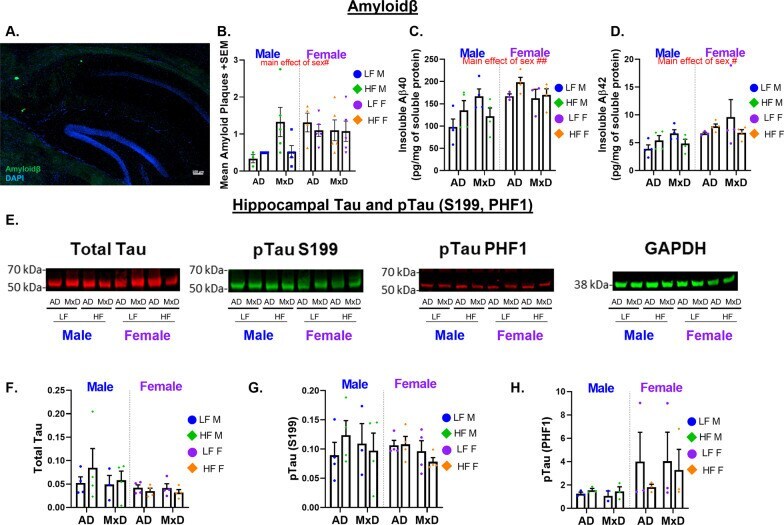
- Female AD/MxD mice have greater Abeta, but not tau, pathology. A Representative image of Abeta plaques. B The number of Abeta plaques/brain section (averaged across 8, 40-muM-thick brain sections between -1.46 and -3.52 mm from bregma). C Hippocampal insoluble Abeta-40, measured by ELISA. D Hippocampal insoluble Abeta-42, measured by ELISA. E Representative Western blot images of tau protein in right (ischemic for MxD) hippocampal isolates. Measurements for F total tau, G pTau S199, and H pTauPHF1 were normalized to GAPDH. Data are presented as mean + SEM, # p < 0.05 effect of sex, ## p < 0.01 effect of sex, 3-way ANOVA, ( n = 4-5/group)
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Fig. 6 Immunoblotting using hTg-Tau and Tau-KO brain lysates confirms the specificity for tau for a panel of antibodies. a ) Scheme of tau (2N4R) showing the epitopes of the Tau12, Tau5, Tau1, HT7, BT2 and Dako antibodies. b ) - d' ) Immunoblots demonstrate strong reactivity of all antibodies with tau bands between 40 and 60 kD. Minor non-specific bands present in Tau-KO brain lysates were detected in d ), e ), i ), h ), p ), x ), y ) and c '). Isoform-specific antibodies in h ), i ), j ) and k ) were validated using a tau ladder with all 6 recombinant human tau isoforms. kDa sizes for the marker are given in e' ). Tg: hTg-Tau mouse, KO: Tau-KO mouse brain lysate, TL: recombinant tau ladder

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
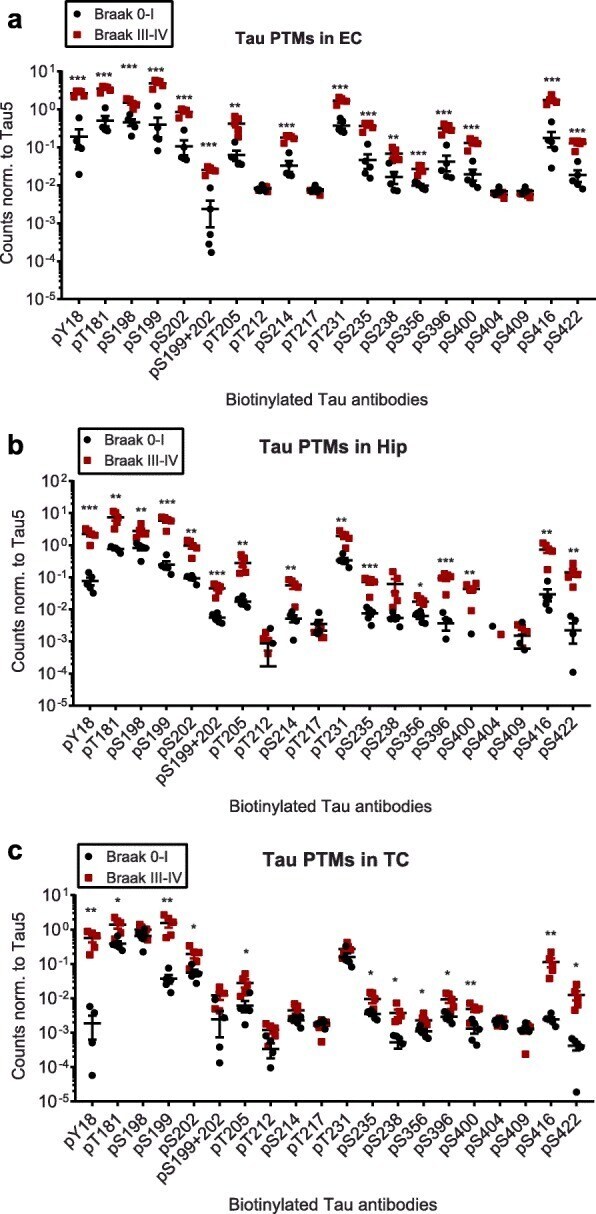
- Fig. 2 Compared to Braak 0-I samples, many but not all, tau phosphorylation events are increased in native Braak III-IV samples. Normalized phospho-tau signals obtained from ELISA measurements of samples from a ) entorhinal cortices (EC), b ) hippocampi (Hip) and c ) temporal cortices (TC). Student's t-tests: *, p < 0.05, **, p < 0.01, ***, p < 0.001