 Explore
Explore Validate
Validate Learn
Learn Western blot
Western blotAntibody data
- Antibody Data
- Antigen structure
- References [75]
- Comments [0]
- Validations
- Western blot [3]
- Immunocytochemistry [5]
- Immunohistochemistry [3]
- Other assay [20]
Submit
Validation data
Reference
Comment
Report error
- Product number
- MA3-006 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- HSP70 Monoclonal Antibody (3A3)
- Antibody type
- Monoclonal
- Antigen
- Recombinant full-length protein
- Description
- MA3-006 detects several members of the heat shock protein 70 kDa (HSP70) gene family including HSP70, HSC70, p75, and following heat shock, HSP72 from yeast, Drosophila, fish, porcine, plant, mouse, avian, arthropod (red claw crayfish & black tiger prawn), amphibian, non-human primate, and human samples.
- Antibody clone number
- 3A3
- Concentration
- 1 mg/mL
Submitted references Nanoscale segregation of channel and barrier claudins enables paracellular ion flux.
Neuroprotective Effects of Asparagus officinalis Stem Extract in Transgenic Mice Overexpressing Amyloid Precursor Protein.
Novel Small Molecule Hsp90/Cdc37 Interface Inhibitors Indirectly Target K-Ras-Signaling.
Basic Limonoid modulates Chaperone-mediated Proteostasis and dissolve Tau fibrils.
Intersectin-Mediated Clearance of SNARE Complexes Is Required for Fast Neurotransmission.
Visualizing tRNA-dependent mistranslation in human cells.
Inhibition of heat shock protein (Hsp) 90 potentiates the antiproliferative and pro-apoptotic effects of 2-(4'fluoro-phenylamino)-4H-1,3-thiazine[6,5-b]indole in A2780cis cells.
Short-term molecular and physiological responses to heat stress in neritic copepods Acartia tonsa and Eurytemora affinis.
Extended Synaptotagmin (ESyt) Triple Knock-Out Mice Are Viable and Fertile without Obvious Endoplasmic Reticulum Dysfunction.
Responses to thermal and salinity stress in wild and farmed Pacific oysters Crassostrea gigas.
Non-lethal heat shock increases tolerance to metal exposure in brine shrimp.
Loss of thermal refugia near equatorial range limits.
Chronic hyperosmotic stress interferes with immune homeostasis in striped catfish (Pangasianodon hypophthalmus, S.) and leads to excessive inflammatory response during bacterial infection.
Non-Lethal Heat Shock of the Asian Green Mussel, Perna viridis, Promotes Hsp70 Synthesis, Induces Thermotolerance and Protects Against Vibrio Infection.
Loss of Gadkin Affects Dendritic Cell Migration In Vitro.
Contrasting roles for MyoD in organizing myogenic promoter structures during embryonic skeletal muscle development.
Histological, ultrastructural and heat shock protein 70 (HSP70) responses to heat stress in the sea cucumber Apostichopus japonicus.
Thermal tolerance in juvenile King George whiting (Sillaginodes punctata) reduces as fish age and this reduction coincides with migration to deeper colder water.
Environmental heat stress induces epigenetic transgenerational inheritance of robustness in parthenogenetic Artemia model.
Non-lethal heat shock increased Hsp70 and immune protein transcripts but not Vibrio tolerance in the white-leg shrimp.
UV-B exposure causes DNA damage and changes in protein expression in northern pike (Esox lucius) posthatched embryos.
The oncogenic TBC domain protein USP6/TRE17 regulates cell migration and cytokinesis.
Enhancement of Hsp70 synthesis protects common carp, Cyprinus carpio L., against lethal ammonia toxicity.
Level of heat shock proteins decreases in individuals carrying B-chromosomes in the grasshopper Eyprepocnemis plorans.
Does domestication process affect stress response in juvenile Eurasian perch Perca fluviatilis?
Priming the prophenoloxidase system of Artemia franciscana by heat shock proteins protects against Vibrio campbellii challenge.
Different rotavirus strains enter MA104 cells through different endocytic pathways: the role of clathrin-mediated endocytosis.
Efficacy of heterologous and homologous heat shock protein 70s as protective agents to Artemia franciscana challenged with Vibrio campbellii.
NF-κB is not directly responsible for photoresistance induced by fractionated light delivery in HT-29 colon adenocarcinoma cells.
Do historical sediments of pulp and paper industry contribute to the exposure of fish caged in receiving waters?
Oxidative stress, protein carbonylation and heat shock proteins in the black tiger shrimp, Penaeus monodon, following exposure to endosulfan and deltamethrin.
The role of p53 in the efficiency of photodynamic therapy with hypericin and subsequent long-term survival of colon cancer cells.
Bioinformatics and protein expression analyses implicate LEA proteins in the drought response of Collembola.
Sex-, gametogenesis, and tidal height-related differences in levels of HSP70 and metallothioneins in the Pacific oyster Crassostrea gigas.
Surface expression of a C-terminal alpha-helix region in heat shock protein 72 on murine LL/2 lung carcinoma can be recognized by innate immune sentinels.
The effects of treated effluents on the intensity of papillomatosis and HSP70 expression in roach.
Saccharomyces cerevisiae strains from traditional fermentations of Brazilian cachaça: trehalose metabolism, heat and ethanol resistance.
Dimeric heat shock protein 40 binds radial spokes for generating coupled power strokes and recovery strokes of 9 + 2 flagella.
Is HSP70 involved in acclimation to cadmium in the Chinese crab, Eriocheir sinensis ?
Heat shock protein (hsp70) expression and thermal tolerance in sublethally heat-shocked eastern oysters Crassostrea virginica infected with the parasite Perkinsus marinus.
Synergistic impacts of heat shock and spawning on the physiology and immune health of Crassostrea gigas: an explanation for summer mortality in Pacific oysters.
Non-lethal heat shock protects gnotobiotic Artemia franciscana larvae against virulent Vibrios.
Thermoprotection of synaptic transmission in a Drosophila heat shock factor mutant is accompanied by increased expression of Hsp83 and DnaJ-1.
Temperature regulates hypoxia-inducible factor-1 (HIF-1) in a poikilothermic vertebrate, crucian carp (Carassius carassius).
Stress protein response in two sibling species of Marenzelleria (Polychaeta: Spionidae): is there an influence of acclimation salinity?
Multiscale approach of fish responses to different types of environmental contaminations: a case study.
Characterization of a molecular chaperone present in the eukaryotic flagellum.
ATP production in Chlamydomonas reinhardtii flagella by glycolytic enzymes.
Dimeric novel HSP40 is incorporated into the radial spoke complex during the assembly process in flagella.
High sensitivity of northern pike larvae to UV-B but no UV-photoinduced toxicity of retene.
A field methodology to study effects of UV radiation on fish larvae.
Peptidoglycan recognition protein tag7 forms a cytotoxic complex with heat shock protein 70 in solution and in lymphocytes.
Rapid glucocorticoid receptor exchange at a promoter is coupled to transcription and regulated by chaperones and proteasomes.
Targeted disruption of the heat shock transcription factor (hsf)-2 gene results in increased embryonic lethality, neuronal defects, and reduced spermatogenesis.
Heat-shock protein (HSP70) response in the eastern oyster, Crassostrea virginica, exposed to PAHs sorbed to suspended artificial clay particles and to suspended field contaminated sediments.
Reconstitution of a functional core polycomb repressive complex.
Co-chaperones Bag-1, Hop and Hsp40 regulate Hsc70 and Hsp90 interactions with wild-type or mutant p53.
Insights into regulation and function of the major stress-induced hsp70 molecular chaperone in vivo: analysis of mice with targeted gene disruption of the hsp70.1 or hsp70.3 gene.
Contribution of heat shock proteins to cell protection from complement-mediated lysis.
Individual subunits of the eukaryotic cytosolic chaperonin mediate interactions with binding sites located on subdomains of beta-actin.
Multiple components of the HSP90 chaperone complex function in regulation of heat shock factor 1 In vivo.
Heat shock response and protein degradation: regulation of HSF2 by the ubiquitin-proteasome pathway.
Effect of different environmental variables on the synthesis of Hsp70 in Raphidocelis subcapitata.
Synthesis and turnover of embryonic sea urchin ciliary proteins during selective inhibition of tubulin synthesis and assembly.
HSP70 binding sites in the tumor suppressor protein p53.
Transport of a novel complex in the cytoplasmic matrix of Chlamydomonas flagella.
Identification of a molecular chaperone in the eukaryotic flagellum and its localization to the site of microtubule assembly.
The RCC1 protein interacts with Ran, RanBP1, hsc70, and a 340-kDa protein in Xenopus extracts.
Cloning and subcellular localization of human mitochondrial hsp70.
Interaction between heat shock factor and hsp70 is insufficient to suppress induction of DNA-binding activity in vivo.
The stress response to loss of signal recognition particle function in Saccharomyces cerevisiae.
A constitutive form of heat-shock protein 70 is located in the outer membranes of mitochondria from rat liver.
Pork quality and the expression of stress protein Hsp 70 in swine.
Regulation of the human hsp70 promoter by p53.
T-complex polypeptide-1 is a subunit of a heteromeric particle in the eukaryotic cytosol.
Gonschior H, Schmied C, Van der Veen RE, Eichhorst J, Himmerkus N, Piontek J, Günzel D, Bleich M, Furuse M, Haucke V, Lehmann M
Nature communications 2022 Aug 25;13(1):4985
Nature communications 2022 Aug 25;13(1):4985
Neuroprotective Effects of Asparagus officinalis Stem Extract in Transgenic Mice Overexpressing Amyloid Precursor Protein.
Peng Z, Bedi S, Mann V, Sundaresan A, Homma K, Gaskey G, Kowada M, Umar S, Kulkarni AD, Eltzschig HK, Doursout MF
Journal of immunology research 2021;2021:8121407
Journal of immunology research 2021;2021:8121407
Novel Small Molecule Hsp90/Cdc37 Interface Inhibitors Indirectly Target K-Ras-Signaling.
Siddiqui FA, Parkkola H, Vukic V, Oetken-Lindholm C, Jaiswal A, Kiriazis A, Pavic K, Aittokallio T, Salminen TA, Abankwa D
Cancers 2021 Feb 23;13(4)
Cancers 2021 Feb 23;13(4)
Basic Limonoid modulates Chaperone-mediated Proteostasis and dissolve Tau fibrils.
Gorantla NV, Das R, Chidambaram H, Dubey T, Mulani FA, Thulasiram HV, Chinnathambi S
Scientific reports 2020 Mar 4;10(1):4023
Scientific reports 2020 Mar 4;10(1):4023
Intersectin-Mediated Clearance of SNARE Complexes Is Required for Fast Neurotransmission.
Jäpel M, Gerth F, Sakaba T, Bacetic J, Yao L, Koo SJ, Maritzen T, Freund C, Haucke V
Cell reports 2020 Jan 14;30(2):409-420.e6
Cell reports 2020 Jan 14;30(2):409-420.e6
Visualizing tRNA-dependent mistranslation in human cells.
Lant JT, Berg MD, Sze DHW, Hoffman KS, Akinpelu IC, Turk MA, Heinemann IU, Duennwald ML, Brandl CJ, O'Donoghue P
RNA biology 2018;15(4-5):567-575
RNA biology 2018;15(4-5):567-575
Inhibition of heat shock protein (Hsp) 90 potentiates the antiproliferative and pro-apoptotic effects of 2-(4'fluoro-phenylamino)-4H-1,3-thiazine[6,5-b]indole in A2780cis cells.
Solárová Z, Kello M, Varinská L, Budovská M, Solár P
Biomedicine & pharmacotherapy = Biomedecine & pharmacotherapie 2017 Jan;85:463-471
Biomedicine & pharmacotherapy = Biomedecine & pharmacotherapie 2017 Jan;85:463-471
Short-term molecular and physiological responses to heat stress in neritic copepods Acartia tonsa and Eurytemora affinis.
Rahlff J, Peters J, Moyano M, Pless O, Claussen C, Peck MA
Comparative biochemistry and physiology. Part A, Molecular & integrative physiology 2017 Jan;203:348-358
Comparative biochemistry and physiology. Part A, Molecular & integrative physiology 2017 Jan;203:348-358
Extended Synaptotagmin (ESyt) Triple Knock-Out Mice Are Viable and Fertile without Obvious Endoplasmic Reticulum Dysfunction.
Sclip A, Bacaj T, Giam LR, Südhof TC
PloS one 2016;11(6):e0158295
PloS one 2016;11(6):e0158295
Responses to thermal and salinity stress in wild and farmed Pacific oysters Crassostrea gigas.
Yang CY, Sierp MT, Abbott CA, Li Y, Qin JG
Comparative biochemistry and physiology. Part A, Molecular & integrative physiology 2016 Nov;201:22-29
Comparative biochemistry and physiology. Part A, Molecular & integrative physiology 2016 Nov;201:22-29
Non-lethal heat shock increases tolerance to metal exposure in brine shrimp.
Pestana JLT, Novais SC, Norouzitallab P, Vandegehuchte MB, Bossier P, De Schamphelaere KAC
Environmental research 2016 Nov;151:663-670
Environmental research 2016 Nov;151:663-670
Loss of thermal refugia near equatorial range limits.
Lima FP, Gomes F, Seabra R, Wethey DS, Seabra MI, Cruz T, Santos AM, Hilbish TJ
Global change biology 2016 Jan;22(1):254-63
Global change biology 2016 Jan;22(1):254-63
Chronic hyperosmotic stress interferes with immune homeostasis in striped catfish (Pangasianodon hypophthalmus, S.) and leads to excessive inflammatory response during bacterial infection.
Schmitz M, Douxfils J, Mandiki SN, Morana C, Baekelandt S, Kestemont P
Fish & shellfish immunology 2016 Aug;55:550-8
Fish & shellfish immunology 2016 Aug;55:550-8
Non-Lethal Heat Shock of the Asian Green Mussel, Perna viridis, Promotes Hsp70 Synthesis, Induces Thermotolerance and Protects Against Vibrio Infection.
Aleng NA, Sung YY, MacRae TH, Abd Wahid ME
PloS one 2015;10(8):e0135603
PloS one 2015;10(8):e0135603
Loss of Gadkin Affects Dendritic Cell Migration In Vitro.
Schachtner H, Weimershaus M, Stache V, Plewa N, Legler DF, Höpken UE, Maritzen T
PloS one 2015;10(12):e0143883
PloS one 2015;10(12):e0143883
Contrasting roles for MyoD in organizing myogenic promoter structures during embryonic skeletal muscle development.
Cho OH, Mallappa C, Hernández-Hernández JM, Rivera-Pérez JA, Imbalzano AN
Developmental dynamics : an official publication of the American Association of Anatomists 2015 Jan;244(1):43-55
Developmental dynamics : an official publication of the American Association of Anatomists 2015 Jan;244(1):43-55
Histological, ultrastructural and heat shock protein 70 (HSP70) responses to heat stress in the sea cucumber Apostichopus japonicus.
Xu D, Sun L, Liu S, Zhang L, Yang H
Fish & shellfish immunology 2015 Aug;45(2):321-6
Fish & shellfish immunology 2015 Aug;45(2):321-6
Thermal tolerance in juvenile King George whiting (Sillaginodes punctata) reduces as fish age and this reduction coincides with migration to deeper colder water.
Meakin CA, Qin JG, Pogson LD, Abbott CA
Comparative biochemistry and physiology. Part A, Molecular & integrative physiology 2014 Jun;172:46-51
Comparative biochemistry and physiology. Part A, Molecular & integrative physiology 2014 Jun;172:46-51
Environmental heat stress induces epigenetic transgenerational inheritance of robustness in parthenogenetic Artemia model.
Norouzitallab P, Baruah K, Vandegehuchte M, Van Stappen G, Catania F, Vanden Bussche J, Vanhaecke L, Sorgeloos P, Bossier P
FASEB journal : official publication of the Federation of American Societies for Experimental Biology 2014 Aug;28(8):3552-63
FASEB journal : official publication of the Federation of American Societies for Experimental Biology 2014 Aug;28(8):3552-63
Non-lethal heat shock increased Hsp70 and immune protein transcripts but not Vibrio tolerance in the white-leg shrimp.
Loc NH, Macrae TH, Musa N, Bin Abdullah MD, Abdul Wahid ME, Sung YY
PloS one 2013;8(9):e73199
PloS one 2013;8(9):e73199
UV-B exposure causes DNA damage and changes in protein expression in northern pike (Esox lucius) posthatched embryos.
Vehniäinen ER, Vähäkangas K, Oikari A
Photochemistry and photobiology 2012 Mar-Apr;88(2):363-70
Photochemistry and photobiology 2012 Mar-Apr;88(2):363-70
The oncogenic TBC domain protein USP6/TRE17 regulates cell migration and cytokinesis.
Rueckert C, Haucke V
Biology of the cell 2012 Jan;104(1):22-33
Biology of the cell 2012 Jan;104(1):22-33
Enhancement of Hsp70 synthesis protects common carp, Cyprinus carpio L., against lethal ammonia toxicity.
Sung YY, Roberts RJ, Bossier P
Journal of fish diseases 2012 Aug;35(8):563-8
Journal of fish diseases 2012 Aug;35(8):563-8
Level of heat shock proteins decreases in individuals carrying B-chromosomes in the grasshopper Eyprepocnemis plorans.
Teruel M, Sørensen JG, Loeschcke V, Cabrero J, Perfectti F, Camacho JP
Cytogenetic and genome research 2011;132(1-2):94-9
Cytogenetic and genome research 2011;132(1-2):94-9
Does domestication process affect stress response in juvenile Eurasian perch Perca fluviatilis?
Douxfils J, Mandiki SN, Marotte G, Wang N, Silvestre F, Milla S, Henrotte E, Vandecan M, Rougeot C, Mélard C, Kestemont P
Comparative biochemistry and physiology. Part A, Molecular & integrative physiology 2011 May;159(1):92-9
Comparative biochemistry and physiology. Part A, Molecular & integrative physiology 2011 May;159(1):92-9
Priming the prophenoloxidase system of Artemia franciscana by heat shock proteins protects against Vibrio campbellii challenge.
Baruah K, Ranjan J, Sorgeloos P, Macrae TH, Bossier P
Fish & shellfish immunology 2011 Jul;31(1):134-41
Fish & shellfish immunology 2011 Jul;31(1):134-41
Different rotavirus strains enter MA104 cells through different endocytic pathways: the role of clathrin-mediated endocytosis.
Gutiérrez M, Isa P, Sánchez-San Martin C, Pérez-Vargas J, Espinosa R, Arias CF, López S
Journal of virology 2010 Sep;84(18):9161-9
Journal of virology 2010 Sep;84(18):9161-9
Efficacy of heterologous and homologous heat shock protein 70s as protective agents to Artemia franciscana challenged with Vibrio campbellii.
Baruah K, Ranjan J, Sorgeloos P, Bossier P
Fish & shellfish immunology 2010 Nov;29(5):733-9
Fish & shellfish immunology 2010 Nov;29(5):733-9
NF-κB is not directly responsible for photoresistance induced by fractionated light delivery in HT-29 colon adenocarcinoma cells.
Kuliková L, Mikeš J, Hýžďalová M, Palumbo G, Fedoročko P
Photochemistry and photobiology 2010 Nov-Dec;86(6):1285-93
Photochemistry and photobiology 2010 Nov-Dec;86(6):1285-93
Do historical sediments of pulp and paper industry contribute to the exposure of fish caged in receiving waters?
Oikari A, Lahti M, Meriläinen P, Afanasyev S, Krasnov A
Journal of environmental monitoring : JEM 2010 May;12(5):1045-54
Journal of environmental monitoring : JEM 2010 May;12(5):1045-54
Oxidative stress, protein carbonylation and heat shock proteins in the black tiger shrimp, Penaeus monodon, following exposure to endosulfan and deltamethrin.
Dorts J, Silvestre F, Tu HT, Tyberghein AE, Phuong NT, Kestemont P
Environmental toxicology and pharmacology 2009 Sep;28(2):302-10
Environmental toxicology and pharmacology 2009 Sep;28(2):302-10
The role of p53 in the efficiency of photodynamic therapy with hypericin and subsequent long-term survival of colon cancer cells.
Mikes J, Koval' J, Jendzelovský R, Sacková V, Uhrinová I, Kello M, Kuliková L, Fedorocko P
Photochemical & photobiological sciences : Official journal of the European Photochemistry Association and the European Society for Photobiology 2009 Nov;8(11):1558-67
Photochemical & photobiological sciences : Official journal of the European Photochemistry Association and the European Society for Photobiology 2009 Nov;8(11):1558-67
Bioinformatics and protein expression analyses implicate LEA proteins in the drought response of Collembola.
Bahrndorff S, Tunnacliffe A, Wise MJ, McGee B, Holmstrup M, Loeschcke V
Journal of insect physiology 2009 Mar;55(3):210-7
Journal of insect physiology 2009 Mar;55(3):210-7
Sex-, gametogenesis, and tidal height-related differences in levels of HSP70 and metallothioneins in the Pacific oyster Crassostrea gigas.
Meistertzheim AL, Lejart M, Le Goïc N, Thébault MT
Comparative biochemistry and physiology. Part A, Molecular & integrative physiology 2009 Feb;152(2):234-9
Comparative biochemistry and physiology. Part A, Molecular & integrative physiology 2009 Feb;152(2):234-9
Surface expression of a C-terminal alpha-helix region in heat shock protein 72 on murine LL/2 lung carcinoma can be recognized by innate immune sentinels.
Tani F, Ohno M, Furukawa Y, Sakamoto M, Masuda S, Kitabatake N
Molecular immunology 2009 Apr;46(7):1326-39
Molecular immunology 2009 Apr;46(7):1326-39
The effects of treated effluents on the intensity of papillomatosis and HSP70 expression in roach.
Korkea-aho TL, Vehniäinen ER, Kukkonen JV, Taskinen J
Ecotoxicology and environmental safety 2008 Jul;70(3):462-8
Ecotoxicology and environmental safety 2008 Jul;70(3):462-8
Saccharomyces cerevisiae strains from traditional fermentations of Brazilian cachaça: trehalose metabolism, heat and ethanol resistance.
Vianna CR, Silva CL, Neves MJ, Rosa CA
Antonie van Leeuwenhoek 2008 Jan-Feb;93(1-2):205-17
Antonie van Leeuwenhoek 2008 Jan-Feb;93(1-2):205-17
Dimeric heat shock protein 40 binds radial spokes for generating coupled power strokes and recovery strokes of 9 + 2 flagella.
Yang C, Owen HA, Yang P
The Journal of cell biology 2008 Jan 28;180(2):403-15
The Journal of cell biology 2008 Jan 28;180(2):403-15
Is HSP70 involved in acclimation to cadmium in the Chinese crab, Eriocheir sinensis ?
Silvestre F, Trausch G, Devos P
Bulletin of environmental contamination and toxicology 2007 Jun;78(6):432-5
Bulletin of environmental contamination and toxicology 2007 Jun;78(6):432-5
Heat shock protein (hsp70) expression and thermal tolerance in sublethally heat-shocked eastern oysters Crassostrea virginica infected with the parasite Perkinsus marinus.
Encomio VG, Chu FL
Diseases of aquatic organisms 2007 Jul 16;76(3):251-60
Diseases of aquatic organisms 2007 Jul 16;76(3):251-60
Synergistic impacts of heat shock and spawning on the physiology and immune health of Crassostrea gigas: an explanation for summer mortality in Pacific oysters.
Li Y, Qin JG, Abbott CA, Li X, Benkendorff K
American journal of physiology. Regulatory, integrative and comparative physiology 2007 Dec;293(6):R2353-62
American journal of physiology. Regulatory, integrative and comparative physiology 2007 Dec;293(6):R2353-62
Non-lethal heat shock protects gnotobiotic Artemia franciscana larvae against virulent Vibrios.
Yik Sung Y, Van Damme EJ, Sorgeloos P, Bossier P
Fish & shellfish immunology 2007 Apr;22(4):318-26
Fish & shellfish immunology 2007 Apr;22(4):318-26
Thermoprotection of synaptic transmission in a Drosophila heat shock factor mutant is accompanied by increased expression of Hsp83 and DnaJ-1.
Neal SJ, Karunanithi S, Best A, So AK, Tanguay RM, Atwood HL, Westwood JT
Physiological genomics 2006 May 16;25(3):493-501
Physiological genomics 2006 May 16;25(3):493-501
Temperature regulates hypoxia-inducible factor-1 (HIF-1) in a poikilothermic vertebrate, crucian carp (Carassius carassius).
Rissanen E, Tranberg HK, Sollid J, Nilsson GE, Nikinmaa M
The Journal of experimental biology 2006 Mar;209(Pt 6):994-1003
The Journal of experimental biology 2006 Mar;209(Pt 6):994-1003
Stress protein response in two sibling species of Marenzelleria (Polychaeta: Spionidae): is there an influence of acclimation salinity?
Blank M, Bastrop R, Jürss K
Comparative biochemistry and physiology. Part B, Biochemistry & molecular biology 2006 Aug;144(4):451-62
Comparative biochemistry and physiology. Part B, Biochemistry & molecular biology 2006 Aug;144(4):451-62
Multiscale approach of fish responses to different types of environmental contaminations: a case study.
Mayon N, Bertrand A, Leroy D, Malbrouck C, Mandiki SN, Silvestre F, Goffart A, Thomé JP, Kestemont P
The Science of the total environment 2006 Aug 31;367(2-3):715-31
The Science of the total environment 2006 Aug 31;367(2-3):715-31
Characterization of a molecular chaperone present in the eukaryotic flagellum.
Shapiro J, Ingram J, Johnson KA
Eukaryotic cell 2005 Sep;4(9):1591-4
Eukaryotic cell 2005 Sep;4(9):1591-4
ATP production in Chlamydomonas reinhardtii flagella by glycolytic enzymes.
Mitchell BF, Pedersen LB, Feely M, Rosenbaum JL, Mitchell DR
Molecular biology of the cell 2005 Oct;16(10):4509-18
Molecular biology of the cell 2005 Oct;16(10):4509-18
Dimeric novel HSP40 is incorporated into the radial spoke complex during the assembly process in flagella.
Yang C, Compton MM, Yang P
Molecular biology of the cell 2005 Feb;16(2):637-48
Molecular biology of the cell 2005 Feb;16(2):637-48
High sensitivity of northern pike larvae to UV-B but no UV-photoinduced toxicity of retene.
Häkkinen J, Vehniäinen E, Oikari A
Aquatic toxicology (Amsterdam, Netherlands) 2004 Mar 10;66(4):393-404
Aquatic toxicology (Amsterdam, Netherlands) 2004 Mar 10;66(4):393-404
A field methodology to study effects of UV radiation on fish larvae.
Häkkinen J, Oikari A
Water research 2004 Jul;38(12):2891-7
Water research 2004 Jul;38(12):2891-7
Peptidoglycan recognition protein tag7 forms a cytotoxic complex with heat shock protein 70 in solution and in lymphocytes.
Sashchenko LP, Dukhanina EA, Yashin DV, Shatalov YV, Romanova EA, Korobko EV, Demin AV, Lukyanova TI, Kabanova OD, Khaidukov SV, Kiselev SL, Gabibov AG, Gnuchev NV, Georgiev GP
The Journal of biological chemistry 2004 Jan 16;279(3):2117-24
The Journal of biological chemistry 2004 Jan 16;279(3):2117-24
Rapid glucocorticoid receptor exchange at a promoter is coupled to transcription and regulated by chaperones and proteasomes.
Stavreva DA, Müller WG, Hager GL, Smith CL, McNally JG
Molecular and cellular biology 2004 Apr;24(7):2682-97
Molecular and cellular biology 2004 Apr;24(7):2682-97
Targeted disruption of the heat shock transcription factor (hsf)-2 gene results in increased embryonic lethality, neuronal defects, and reduced spermatogenesis.
Wang G, Zhang J, Moskophidis D, Mivechi NF
Genesis (New York, N.Y. : 2000) 2003 May;36(1):48-61
Genesis (New York, N.Y. : 2000) 2003 May;36(1):48-61
Heat-shock protein (HSP70) response in the eastern oyster, Crassostrea virginica, exposed to PAHs sorbed to suspended artificial clay particles and to suspended field contaminated sediments.
Cruz-Rodríguez LA, Chu FL
Aquatic toxicology (Amsterdam, Netherlands) 2002 Oct 30;60(3-4):157-68
Aquatic toxicology (Amsterdam, Netherlands) 2002 Oct 30;60(3-4):157-68
Reconstitution of a functional core polycomb repressive complex.
Francis NJ, Saurin AJ, Shao Z, Kingston RE
Molecular cell 2001 Sep;8(3):545-56
Molecular cell 2001 Sep;8(3):545-56
Co-chaperones Bag-1, Hop and Hsp40 regulate Hsc70 and Hsp90 interactions with wild-type or mutant p53.
King FW, Wawrzynow A, Höhfeld J, Zylicz M
The EMBO journal 2001 Nov 15;20(22):6297-305
The EMBO journal 2001 Nov 15;20(22):6297-305
Insights into regulation and function of the major stress-induced hsp70 molecular chaperone in vivo: analysis of mice with targeted gene disruption of the hsp70.1 or hsp70.3 gene.
Huang L, Mivechi NF, Moskophidis D
Molecular and cellular biology 2001 Dec;21(24):8575-91
Molecular and cellular biology 2001 Dec;21(24):8575-91
Contribution of heat shock proteins to cell protection from complement-mediated lysis.
Fishelson Z, Hochman I, Greene LE, Eisenberg E
International immunology 2001 Aug;13(8):983-91
International immunology 2001 Aug;13(8):983-91
Individual subunits of the eukaryotic cytosolic chaperonin mediate interactions with binding sites located on subdomains of beta-actin.
Hynes GM, Willison KR
The Journal of biological chemistry 2000 Jun 23;275(25):18985-94
The Journal of biological chemistry 2000 Jun 23;275(25):18985-94
Multiple components of the HSP90 chaperone complex function in regulation of heat shock factor 1 In vivo.
Bharadwaj S, Ali A, Ovsenek N
Molecular and cellular biology 1999 Dec;19(12):8033-41
Molecular and cellular biology 1999 Dec;19(12):8033-41
Heat shock response and protein degradation: regulation of HSF2 by the ubiquitin-proteasome pathway.
Mathew A, Mathur SK, Morimoto RI
Molecular and cellular biology 1998 Sep;18(9):5091-8
Molecular and cellular biology 1998 Sep;18(9):5091-8
Effect of different environmental variables on the synthesis of Hsp70 in Raphidocelis subcapitata.
Bierkens J, Van de Perre WV, Maes J
Comparative biochemistry and physiology. Part A, Molecular & integrative physiology 1998 May;120(1):29-34
Comparative biochemistry and physiology. Part A, Molecular & integrative physiology 1998 May;120(1):29-34
Synthesis and turnover of embryonic sea urchin ciliary proteins during selective inhibition of tubulin synthesis and assembly.
Stephens RE
Molecular biology of the cell 1997 Nov;8(11):2187-98
Molecular biology of the cell 1997 Nov;8(11):2187-98
HSP70 binding sites in the tumor suppressor protein p53.
Fourie AM, Hupp TR, Lane DP, Sang BC, Barbosa MS, Sambrook JF, Gething MJ
The Journal of biological chemistry 1997 Aug 1;272(31):19471-9
The Journal of biological chemistry 1997 Aug 1;272(31):19471-9
Transport of a novel complex in the cytoplasmic matrix of Chlamydomonas flagella.
Piperno G, Mead K
Proceedings of the National Academy of Sciences of the United States of America 1997 Apr 29;94(9):4457-62
Proceedings of the National Academy of Sciences of the United States of America 1997 Apr 29;94(9):4457-62
Identification of a molecular chaperone in the eukaryotic flagellum and its localization to the site of microtubule assembly.
Bloch MA, Johnson KA
Journal of cell science 1995 Nov;108 ( Pt 11):3541-5
Journal of cell science 1995 Nov;108 ( Pt 11):3541-5
The RCC1 protein interacts with Ran, RanBP1, hsc70, and a 340-kDa protein in Xenopus extracts.
Saitoh H, Dasso M
The Journal of biological chemistry 1995 May 5;270(18):10658-63
The Journal of biological chemistry 1995 May 5;270(18):10658-63
Cloning and subcellular localization of human mitochondrial hsp70.
Bhattacharyya T, Karnezis AN, Murphy SP, Hoang T, Freeman BC, Phillips B, Morimoto RI
The Journal of biological chemistry 1995 Jan 27;270(4):1705-10
The Journal of biological chemistry 1995 Jan 27;270(4):1705-10
Interaction between heat shock factor and hsp70 is insufficient to suppress induction of DNA-binding activity in vivo.
Rabindran SK, Wisniewski J, Li L, Li GC, Wu C
Molecular and cellular biology 1994 Oct;14(10):6552-60
Molecular and cellular biology 1994 Oct;14(10):6552-60
The stress response to loss of signal recognition particle function in Saccharomyces cerevisiae.
Arnold CE, Wittrup KD
The Journal of biological chemistry 1994 Dec 2;269(48):30412-8
The Journal of biological chemistry 1994 Dec 2;269(48):30412-8
A constitutive form of heat-shock protein 70 is located in the outer membranes of mitochondria from rat liver.
Lithgow T, Ryan M, Anderson RL, Høj PB, Hoogenraad NJ
FEBS letters 1993 Oct 18;332(3):277-81
FEBS letters 1993 Oct 18;332(3):277-81
Pork quality and the expression of stress protein Hsp 70 in swine.
van Laack RL, Faustman C, Sebranek JG
Journal of animal science 1993 Nov;71(11):2958-64
Journal of animal science 1993 Nov;71(11):2958-64
Regulation of the human hsp70 promoter by p53.
Agoff SN, Hou J, Linzer DI, Wu B
Science (New York, N.Y.) 1993 Jan 1;259(5091):84-7
Science (New York, N.Y.) 1993 Jan 1;259(5091):84-7
T-complex polypeptide-1 is a subunit of a heteromeric particle in the eukaryotic cytosol.
Lewis VA, Hynes GM, Zheng D, Saibil H, Willison K
Nature 1992 Jul 16;358(6383):249-52
Nature 1992 Jul 16;358(6383):249-52
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
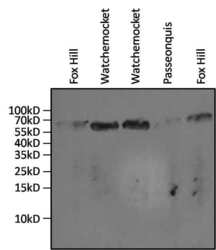
- Experimental details
- Western blot analysis of Hsp70 was performed by loading 20 µL of gill tissue lysates from the salt marsh mussel, Guekensia demissa, isolated from various coves in Rhode Island (indicated above the lanes) and processed by homogenization in 32mM Tris-HCl, 1mM EDTA, and 2% SDS homogenization buffer, per well onto an SDS-PAGE gel. Proteins extracted from the supernatants were transferred to nitrocellulose membrane and blocked overnight with 5% milk in TBST. The membrane was probed with an Hsp70 monoclonal antibody (Product # MA3-006) at a dilution of 1:1000, washed in TBST, and probed with an HRP-conjugated goat anti-mouse IgG secondary antibody at a dilution of 1:3000. Detection was performed using SuperSignal West Pico (Product # 34080). Data courtesy of the Innovators Program.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Western blot was performed using Anti-HSP70 Monoclonal Antibody (3A3) (Product # MA3-006) and a 70kDa band corresponding to HSP70 was observed across cell lines and tissues tested and was induced upon heat shock in Jurkat cell line. Whole cell extracts (30 µg lysate) of HEK-293 (Lane 1), A549 (Lane 2), IMR-32 (Lane 3), Jurkat (Lane 4), Jurkat treated with heat shock (42deg for 30 min followed by 37deg for 3 hours) (Lane 5), MDCK (Lane 6), NIH/3T3 (Lane 7), PC-12 (Lane 8), COS-7 (Lane 9) and Mouse Ovary (Lane 10) were electrophoresed using Novex® NuPAGE® 4-12 % Bis-Tris gel (Product # NP0322BOX). Resolved proteins were then transferred onto a nitrocellulose membrane (Product # IB23001) by iBlot® 2 Dry Blotting System (Product # IB21001). The blot was probed with the primary antibody (1:1000 dilution) and detected by chemiluminescence with Goat anti-Mouse IgG (H+L) Superclonal™ Recombinant Secondary Antibody, HRP (Product # A28177, 1:4000 dilution) using the iBright FL 1000 (Product # A32752). Chemiluminescent detection was performed using Novex® ECL Chemiluminescent Substrate Reagent Kit (Product # WP20005).

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Western blot analysis of Heat Shock Protein 70 (Hsp70) was performed by loading 50 µg of the indicated whole cell lysates and 15 µL of PageRuler Prestained Protein Ladder (Product # 26616) onto a 4-20% Tris-HCl polyacrylamide gel. Proteins were transferred to a PVDF membrane and blocked with 5% BSA/TBST for at least 1 hour. The membrane was probed with a Hsp70 monoclonal antibody (Product # MA3-006) at a dilution of 1:1000 overnight at 4°C on a rocking platform, washed in TBS-0.1%Tween 20, and probed with a goat anti-mouse IgG-HRP secondary antibody (Product # 31430) at a dilution of 1:20,000 for at least 1 hour. Chemiluminescent detection was performed using SuperSignal West Pico (Product # 34080).

Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
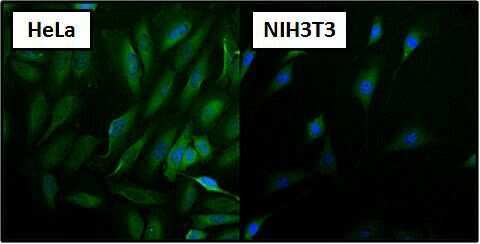
- Immunofluorescent analysis of Heat Shock Protein 70 using Heat Shock Protein 70 Monoclonal antibody (3A3) (Product # MA3-006) shows staining in NIH-3T3 cells. Heat Shock Protein 70 staining (green), F-Actin staining with Phalloidin (red) and nuclei with DAPI (blue) is shown. Cells were grown on chamber slides and fixed with formaldehyde prior to staining. Cells were probed without (control) or with or an antibody recognizing Heat Shock Protein 70 (Product # MA3-006) at a dilution of 1:100-1:200 over night at 4 °C, washed with PBS and incubated with a DyLight-488 conjugated secondary antibody (Product # 35552 for GAR, Product # 35503 for GAM). Images were taken at 60X magnification.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
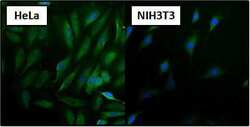
- Immunofluorescent analysis of Heat Shock Protein 70 using Heat Shock Protein 70 Monoclonal antibody (3A3) (Product # MA3-006) shows staining in HeLa cells. Heat Shock Protein 70 staining (green), F-Actin staining with Phalloidin (red) and nuclei with DAPI (blue) is shown. Cells were grown on chamber slides and fixed with formaldehyde prior to staining. Cells were probed without (control) or with or an antibody recognizing Heat Shock Protein 70 (Product # MA3-006) at a dilution of 1:100-1:200 over night at 4 °C, washed with PBS and incubated with a DyLight-488 conjugated secondary antibody (Product # 35552 for GAR, Product # 35503 for GAM). Images were taken at 60X magnification.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Immunofluorescent analysis of Heat Shock Protein 70 using Heat Shock Protein 70 Monoclonal antibody (3A3) (Product # MA3-006) shows staining in MCF-7 cells. Heat Shock Protein 70 staining (green), F-Actin staining with Phalloidin (red) and nuclei with DAPI (blue) is shown. Cells were grown on chamber slides and fixed with formaldehyde prior to staining. Cells were probed without (control) or with or an antibody recognizing Heat Shock Protein 70 (Product # MA3-006) at a dilution of 1:100-1:200 over night at 4 °C, washed with PBS and incubated with a DyLight-488 conjugated secondary antibody (Product # 35552 for GAR, Product # 35503 for GAM). Images were taken at 60X magnification.
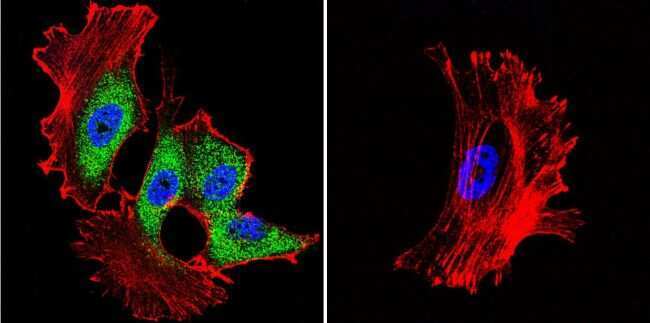
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- Immunofluorescent analysis of Heat Shock Protein 70 (Hsp70) (green) in HeLa and NIH3T3 cells. Formalin fixed cells were permeabilized with 0.1% Triton X-100 in TBS for 10 minutes at room temperature and blocked with 1% Blocker BSA (Product # 37525) for 15 minutes at room temperature. Cells were probed with a Hsp70 monoclonal antibody (Product # MA3-006), at a dilution of 1:50 for at least 1 hour at room temperature, washed with PBS, and incubated with DyLight 488 goat-anti-mouse IgG secondary antibody (Product # 35502) at a dilution of 1:400 for 30 minutes at room temperature. Nuclei (blue) were stained with Hoechst 33342 dye (Product # 62249). Images were taken on a Thermo Scientific ArrayScan at 20X magnification.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Immunofluorescence analysis of HSP70 was performed using 70% confluent log phase HEK-293 cells. The cells were fixed with 4% paraformaldehyde for 10 minutes, permeabilized with 0.1% Triton™ X-100 for 15 minutes, and blocked with 2% BSA for 1 hour at room temperature. The cells were labeled with HSP70 Monoclonal Antibody (3A3) (Product # MA3-006) at 1:200 dilution in 0.1% BSA, incubated at 4 degree Celsius overnight and then labeled with Goat anti-Mouse IgG (H+L), Superclonal™ Recombinant Secondary Antibody, Alexa Fluor 488 conjugate (Product # A28175) at a dilution of 1:2000 for 45 minutes at room temperature (Panel a: green). Nuclei (Panel b: blue) were stained with SlowFade® Gold Antifade Mountant with DAPI (Product # S36938). F-actin (Panel c: red) was stained with Rhodamine Phalloidin (Product # R415, 1:300). Panel d represents the merged image showing localization to nucleus and cytoplasm. Panel e represents control cells with no primary antibody to assess background. The images were captured at 60X magnification.

Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Immunohistochemistry was performed on normal deparaffinized human Testis tissue. To expose target proteins, heat induced antigen retrieval was performed using 10mM sodium citrate (pH6.0) buffer, microwaved for 8-15 minutes. Following antigen retrieval tissues were blocked in 3% BSA-PBS for 30 minutes at room temperature. Tissues were then probed at a dilution of 1:200 with a mouse monoclonal antibody recognizing Heat Shock Protein 70 (Product # MA3-006) or without primary antibody (negative control) overnight at 4°C in a humidified chamber. Tissues were washed extensively with PBST and endogenous peroxidase activity was quenched with a peroxidase suppressor. Detection was performed using a biotin-conjugated secondary antibody and SA-HRP, followed by colorimetric detection using DAB. Tissues were counterstained with hematoxylin and prepped for mounting.
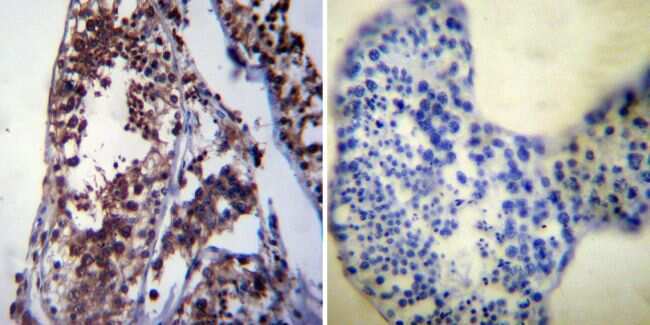
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Immunohistochemistry was performed on normal deparaffinized human Tonsil tissue. To expose target proteins, heat induced antigen retrieval was performed using 10mM sodium citrate (pH6.0) buffer, microwaved for 8-15 minutes. Following antigen retrieval tissues were blocked in 3% BSA-PBS for 30 minutes at room temperature. Tissues were then probed at a dilution of 1:20 with a mouse monoclonal antibody recognizing Heat Shock Protein 70 (Product # MA3-006) or without primary antibody (negative control) overnight at 4°C in a humidified chamber. Tissues were washed extensively with PBST and endogenous peroxidase activity was quenched with a peroxidase suppressor. Detection was performed using a biotin-conjugated secondary antibody and SA-HRP, followed by colorimetric detection using DAB. Tissues were counterstained with hematoxylin and prepped for mounting.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Immunohistochemistry was performed on cancer biopsies of deparaffinized human Breast carcinoma tissue. To expose target proteins, heat induced antigen retrieval was performed using 10mM sodium citrate (pH6.0) buffer, microwaved for 8-15 minutes. Following antigen retrieval tissues were blocked in 3% BSA-PBS for 30 minutes at room temperature. Tissues were then probed at a dilution of 1:50 with a mouse monoclonal antibody recognizing Heat Shock Protein 70 (Product # MA3-006) or without primary antibody (negative control) overnight at 4°C in a humidified chamber. Tissues were washed extensively with PBST and endogenous peroxidase activity was quenched with a peroxidase suppressor. Detection was performed using a biotin-conjugated secondary antibody and SA-HRP, followed by colorimetric detection using DAB. Tissues were counterstained with hematoxylin and prepped for mounting.
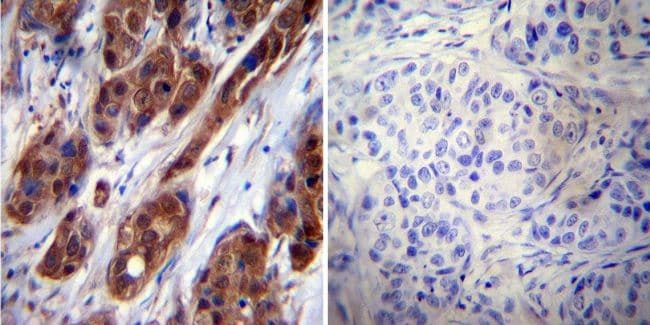
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
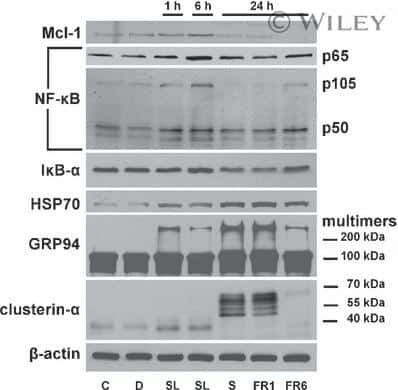
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
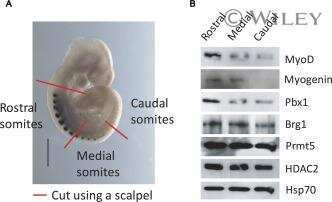
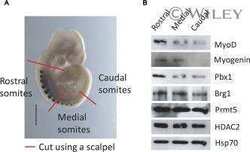
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Fig 2 Hsp70 was induced in P . viridis by heat shock. Protein samples from the adductor muscle (A, B), foot (C, D), gill (E, F) and mantel (G, H) were resolved in 7% SDS polyacrylamide gels and either stained with Coomassie blue (A, C, E, G) or blotted to membranes and probed with antibody to Hsp70 (B, D, F, H). Arrows labeled p70 indicate the position of 70 kDa proteins in the gel. M, molecular mass markers in kDa; H-Hsp70, recombinant human Hsp70.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

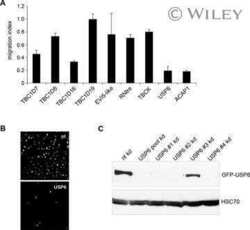
- Experimental details
- Fig 1 Gadkin is expressed in lymphoid organs and in immune cells. A. Lysates of WT and Gadkin -/- (labeled as KO) tissues were analyzed by immunoblotting using Gadkin- and AP-1-specific antibodies. B. Lysates of WT and Gadkin -/- thymus, of DCs derived from WT and Gadkin -/- bone marrow, as well as of B lymphocytes (i.e. B220-positive B cells) and T lymphocytes (i.e. CD3-positive T cells) isolated from spleen were analyzed by immunoblotting using Gadkin-specific antibodies. Detection of the constitutively expressed molecular chaperone heat shock cognate protein 70 (Hsc70) was used as loading control. B, B-lymphocytes; T, T-lymphocytes.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 3 NLHS increased Hsp70 in L. vannamei post-larvae. Protein extracts were resolved by electrophoresis in SDS polyacrylamide gels and either stained with Coomassie Biosafe (A) or blotted to polyvinylidene fluoride membranes and incubated with antibody to Hsp70 (B). Approximately 50 ug of protein was loaded in each lane. 28, non-heat shocked post-larvae heat shock at 30degC, 32degC, 34degC, 36degC and 38degC was for 30 min with recovery at 28degC for 8 h H-Hsp70: Human Hsp70 recombinant protein M: molecular mass standards in kDa. The figure is a representation from two separate experiments.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 4 NLHS enhanced Hsp70 production in L. vannamei . Post-larvae were exposed to 30 min heat shock from 28degC to 30degC, 32degC, 34degC, 36degC and 38degC, then transferred to 28degC for 8 h. Quantification of Hsp70 was by ELISA. A standard curve, constructed with Human Hsp70 recombinant protein was used to convert ELISA readings obtained with shrimp tissue protein extracts to Hsp70 content. Bars represent the fold difference of Hsp70 quantity in comparison to a non-heated control. The error bars represent the SD from 3 replicates. Asterisks denote statistically significant differences between values obtained for control and heat shocked post-larvae ( P< 0.05). The figure is a representation from two separate experiments.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Fig 4 P . viridis Hsp70 Varied with Recovery Time after Heat Shock. Protein samples from the adductor muscle (A, B), foot (C, D), gill (E, F) and mantel (G, H) were resolved in 7% SDS polyacrylamide gels and either stained with Coomassie blue (A, C, E, G) or blotted to membranes and probed with antibody to Hsp70 (B, D, F, H). Arrows labeled p70 indicate the position of 70 kDa proteins in the gel. M, molecular mass markers in kDa; H-Hsp70, recombinant human Hsp70. C, mussels not receiving heat shock.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
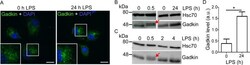
- Experimental details
- Fig 4 Gadkin is regulated in an LPS-dependent manner. A-D. LPS treatment of BMDCs increases Gadkin levels and induces its transient posttranscriptional modification. A. Immature WT BMDCs were either left untreated or incubated for 24 h with LPS. Subsequently, cells were allowed to spread on fibronectin-coated cover slips for 15 min to achieve a more uniform morphology prior to analysis. Cells were processed for immunofluorescence and labeled with Gadkin-specific antibodies. Nuclei were stained with DAPI. Insets show 2x enlarged perinuclear area. Scale bar: 10 mum. B,C. Lysates from WT BMDCs treated with LPS for the indicated times were analyzed by immunoblotting using Gadkin- and Hsc70-specific antibodies. The closely spaced upper band is indicated by red arrows. D. Quantification of immunoblot images like the one depicted in B. Gadkin levels were normalized to Hsc70 levels derived from the same blot. (N = 3 independent experiments, unpaired two-tailed t-test; * = p

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
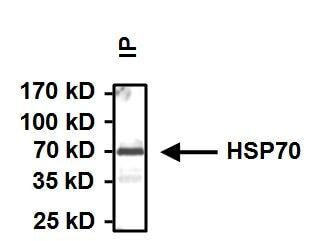
- Immunoprecipitation of Heat Shock Protein 70 (Hsp70) was performed on HeLa cells. Antigen:antibody complexes were formed by incubating 500 µg whole cell lysate with 2 µg of Hsp70 monoclonal antibody (Product # MA3-006) overnight on a rocking platform at 4°C. The immune complexes were captured on 50 µL Protein A/G Plus Agarose (Product # 20421), washed extensively, and eluted with Lane Marker Reducing Sample Buffer (Product # 39000). Samples were then resolved on a 4-20% Tris-HCl polyacrylamide gel, transferred to a PVDF membrane, and blocked with 5% BSA/TBST for at least 1 hour. The membrane was probed with a Hsp70 monoclonal antibody (Product # MA3-006) at a dilution of 1:1000 overnight rotating at 4°C, washed in TBST, and probed with goat anti-mouse IgG-HRP secondary antibody (Product # 31430) at a dilution of 1:20,000 for at least 1 hour. Chemiluminescent detection was performed using SuperSignal West Dura (Product # 34075).
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 6 Top compounds decrease the expression of Hsp90 clients, and dose-dependently inhibit K-Ras nanoclustering-FRET. ( A - D ) Western blots of HER-2 overexpressing SK-BR-3 ( A ), KRAS mutant triple negative breast MDA-MB-231 ( B ), KRAS mutant pancreatic MIA PaCa-2 ( C ), and KRAS mutant lung HCC-44 ( D ) cancer cell lines. The cells were treated with 0.1% DMSO vehicle control or the indicated concentration of compounds. In total, 2 muM 17-AAG or 10 muM conglobatin A served as the positive control. Note the increase in Hsp90 and, in particular, Hsp70 with the 17-AAG treatment, which was not observed with interface inhibitors. All of the drug treatments were for 24 h; n = 1 independent biological repeat. ( E , F ) K-RasG12V- ( E ) and HRasG12V- ( F ) nanoclustering-FRET in HEK cells co-transfected with mGFP- and mCherry-tagged RasG12V. The cells were treated for 24 h with 0.1% DMSO vehicle control, 2 muM conglobatin A, 2 muM compactin, 0.5 muM FTI277, or the indicated concentrations of x6505 or x1540 ( E ) or 5 muM of these hit compounds ( F ). Compactin and FTI277 block Ras prenylation and farnesylation, respectively, and served as positive controls. The numbers in the bars indicate the number of analyzed cells; n = 3 independent biological repeats. **** p < 0.0001.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 3 Quantification of heat shock protein (HSP27) and HSP70 in the hippocampus of APP-overexpressed mice treated with ETAS (200 mg/kg and 1000 mg/kg) and WT mice treated with ETAS (1000 mg/kg) for 1 month, respectively; RFUs: relative fluorescence units; green represents HSP27/70 expression, and blue represents DAPI. Arrows represent fluorescent green staining, indicating positive expression. Magnification is represented x200. HSP27: ** p < 0.01 APP-ETAS (1000 mg/kg) vs. APP-ETAS (200 mg/kg) and APP saline-treated mice; ** p < 0.01 WT-ETAS (1000 mg/kg) vs. WT saline-treated mice; RFUs: relative fluorescence units. Four mice per group and four sections per mouse were used. HSP70: * p < 0.05 APP-ETAS (1000 mg/kg) vs. APP-ETAS (200 mg/kg); ** p < 0.01 APP-ETAS (1000 mg/kg) vs. APP saline-treated mice; ** p < 0.01 WT-ETAS (1000 mg/kg) vs. WT saline-treated mice. RFUs: relative fluorescence units. Four mice per group and four sections per mouse were used.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Fig 6 Two 70 kDa protein bands were resolved in 5% SDS polyacrylamide gels. Tissues corresponding to the adductor muscle, foot, gill and mantel were isolated from P . viridis exposed to NLHS, homogenized, centrifuged and applied to a 5% SDS polyacrylamide gel. Labeled arrows indicate excised regions of the gel subsequently shown by mass spectrometry to contain Pv Hsp70-1 and Pv Hsp70-2. M, molecular mass in kDa; C, mussels not receiving NLHS; HS, mussels receiving NLHS; F, foot; G, gill; MA, mantle; MU, adductor muscle.
