 Explore
Explore Validate
Validate Learn
Learn Flow cytometry
Flow cytometryAntibody data
- Antibody Data
- Antigen structure
- References [85]
- Comments [0]
- Validations
- Flow cytometry [1]
- Other assay [52]
Submit
Validation data
Reference
Comment
Report error
- Product number
- 12-1057-42 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- CD105 (Endoglin) Monoclonal Antibody (SN6), PE, eBioscience™
- Antibody type
- Monoclonal
- Antigen
- Other
- Description
- Description: The SN6 monoclonal antibody reacts with human CD105, also known as Endoglin. CD105, an approximately 90 kDa disulfide-linked homodimer is expressed by vascular endothelial cells and some bone marrow cells and activated macrophages. It is suggested that CD105 functions in adhesion and embryonic angiogenesis. Applications Reported: The SN6 antibody has been reported for use in flow cytometric analysis. Applications Tested: The SN6 antibody has been pre-titrated and tested by flow cytometric analysis of the U937 cell line. This can be used at 5 µL (1 µg) per test. A test is defined as the amount (µg) of antibody that will stain a cell sample in a final volume of 100 µL. Cell number should be determined empirically but can range from 10^5 to 10^8 cells/test. Excitation: 488-561 nm; Emission: 578 nm; Laser: Blue Laser, Green Laser, Yellow-Green Laser. Filtration: 0.2 µm post-manufacturing filtered.
- Reactivity
- Human
- Host
- Mouse
- Conjugate
- Yellow dye
- Isotype
- IgG
- Antibody clone number
- SN6
- Vial size
- 100 Tests
- Concentration
- 5 µL/Test
- Storage
- 4° C, store in dark, DO NOT FREEZE!
Submitted references Single-cell analysis of immune cells on gingiva-derived mesenchymal stem cells in experimental autoimmune uveitis.
Neuronal Cell Differentiation of Human Dental Pulp Stem Cells on Synthetic Polymeric Surfaces Coated With ECM Proteins.
Mesenchymal stem cells derived from patients with premature aging syndromes display hallmarks of physiological aging.
Carboxymethyl chitin or chitosan for osteoinduction effect on the human periodontal ligament stem cells.
Exosomes derived from stem cells of human deciduous exfoliated teeth inhibit angiogenesis in vivo and in vitro via the transfer of miR-100-5p and miR-1246.
Mechanical Compression by Simulating Orthodontic Tooth Movement in an In Vitro Model Modulates Phosphorylation of AKT and MAPKs via TLR4 in Human Periodontal Ligament Cells.
In-cytoplasm mitochondrial transplantation for mesenchymal stem cells engineering and tissue regeneration.
Induced Neurodifferentiation of hBM-MSCs through Activation of the ERK/CREB Pathway via Pulsed Electromagnetic Fields and Physical Stimulation Promotes Neurogenesis in Cerebral Ischemic Models.
Isolation and characterization mesenchymal stem cells from red panda (Ailurus fulgens styani) endometrium.
Production enhancement of human adipose-derived mesenchymal stem cells by low-intensity ultrasound stimulation.
Aberrant Expression of COX-2 and FOXG1 in Infrapatellar Fat Pad-Derived ASCs from Pre-Diabetic Donors.
Human Amniotic Mesenchymal Stem Cells Inhibit aGVHD by Regulating Balance of Treg and T Effector Cells.
Development, characterization, and hematopoietic differentiation of Griscelli syndrome type 2 induced pluripotent stem cells.
Therapeutic potential of human umbilical cord mesenchymal stem cells on aortic atherosclerotic plaque in a high-fat diet rabbit model.
Glycemic control by umbilical cord-derived mesenchymal stem cells promotes effects of fasting-mimicking diet on type 2 diabetic mice.
Differentiation potential and mRNA profiles of human dedifferentiated adipose cells and adipose‑derived stem cells from young donors.
TRPA1 triggers hyperalgesia and inflammation after tooth bleaching.
The Secretome Derived From Mesenchymal Stromal Cells Cultured in a Xeno-Free Medium Promotes Human Cartilage Recovery in vitro.
Down-Regulated Exosomal MicroRNA-221 - 3p Derived From Senescent Mesenchymal Stem Cells Impairs Heart Repair.
Differentiation Potential of Early- and Late-Passage Adipose-Derived Mesenchymal Stem Cells Cultured under Hypoxia and Normoxia.
Vasculogenesis from Human Dental Pulp Stem Cells Grown in Matrigel with Fully Defined Serum-Free Culture Media.
Apoptotic bodies derived from mesenchymal stem cells promote cutaneous wound healing via regulating the functions of macrophages.
The Effect of Angiotensin II, Retinoic Acid, EGCG, and Vitamin C on the Cardiomyogenic Differentiation Induction of Human Amniotic Fluid-Derived Mesenchymal Stem Cells.
Resveratrol rescues TNF‑α‑induced inhibition of osteogenesis in human periodontal ligament stem cells via the ERK1/2 pathway.
Migration Inhibitory Factor in Conditioned Medium from Human Umbilical Cord Blood-Derived Mesenchymal Stromal Cells Stimulates Hair Growth.
Chondrogenic Differentiation from Induced Pluripotent Stem Cells Using Non-Viral Minicircle Vectors.
ZKSCAN3 counteracts cellular senescence by stabilizing heterochromatin.
Rescue of premature aging defects in Cockayne syndrome stem cells by CRISPR/Cas9-mediated gene correction.
Impairment of endothelial progenitor cells function in patient with mustard gas intoxication.
PF-127 hydrogel plus sodium ascorbyl phosphate improves Wharton's jelly mesenchymal stem cell-mediated skin wound healing in mice.
Therapeutic Effects of Human Urine-Derived Stem Cells in a Rat Model of Cisplatin-Induced Acute Kidney Injury In Vivo and In Vitro.
Investigating the potential of the secretome of mesenchymal stem cells derived from sickle cell disease patients.
Ultrastructural morphology is distinct among primary progenitor cell isolates from normal, inflamed, and cryopreserved equine hoof tissue and CD105(+)K14(+) progenitor cells.
Telomere-dependent and telomere-independent roles of RAP1 in regulating human stem cell homeostasis.
TGF‑β induces periodontal ligament stem cell senescence through increase of ROS production.
High-yield isolation of menstrual blood-derived endometrial stem cells by direct red blood cell lysis treatment.
Better therapeutic potential of bone marrow-derived mesenchymal stem cells compared with chorionic villi-derived mesenchymal stem cells in airway injury model.
Stabilizing heterochromatin by DGCR8 alleviates senescence and osteoarthritis.
Bone mesenchymal stromal cells exhibit functional inhibition but no chromosomal aberrations in chronic myelogenous leukemia.
Visfatin Mediates Malignant Behaviors through Adipose-Derived Stem Cells Intermediary in Breast Cancer.
Up-regulation of FOXD1 by YAP alleviates senescence and osteoarthritis.
ATF6 safeguards organelle homeostasis and cellular aging in human mesenchymal stem cells.
The ICAM-1 expression level determines the susceptibility of human endothelial cells to simulated microgravity.
BNIP3/Bcl-2-mediated apoptosis induced by cyclic tensile stretch in human cartilage endplate-derived stem cells.
Isolation and characterization of stem cells from differentially degenerated human lumbar zygapophyseal articular cartilage.
Human umbilical cord-derived mesenchymal stem cells ameliorate the enteropathy of food allergies in mice.
Antagonizing CD105 enhances radiation sensitivity in prostate cancer.
Differential stem cell aging kinetics in Hutchinson-Gilford progeria syndrome and Werner syndrome.
Effects of acute exposure to low-dose radiation on the characteristics of human bone marrow mesenchymal stromal/stem cells.
Insulin-Like Growth Factor Binding Protein-6 Alters Skeletal Muscle Differentiation of Human Mesenchymal Stem Cells.
Endoglin: a novel target for therapeutic intervention in acute leukemias revealed in xenograft mouse models.
A high-yield isolation and enrichment strategy for human lung microvascular endothelial cells.
The Improvement of Respiratory Performance After Phototherapy-Induced EPC Mobilization in Preterm Infants With RDS.
Human umbilical cord mesenchymal stem cells improve the reserve function of perimenopausal ovary via a paracrine mechanism.
Elimination of undifferentiated human embryonic stem cells by cardiac glycosides.
Circulation Enrichment of Functional Endothelial Progenitor Cells by Infantile Phototherapy.
Feasibility of placenta-derived mesenchymal stem cells as a tool for studying pregnancy-related disorders.
A Member of the Nuclear Receptor Superfamily, Designated as NR2F2, Supports the Self-Renewal Capacity and Pluripotency of Human Bone Marrow-Derived Mesenchymal Stem Cells.
CRISPR/Cas9-based genetic correction for recessive dystrophic epidermolysis bullosa.
Influence of aging on the quantity and quality of human cardiac stem cells.
Angiogenic activity mediates bone repair from human pluripotent stem cell-derived osteogenic cells.
Potency of umbilical cord blood- and Wharton's jelly-derived mesenchymal stem cells for scarless wound healing.
SIRT6 safeguards human mesenchymal stem cells from oxidative stress by coactivating NRF2.
Lymphatic Contribution to the Cellular Niche in Heterotopic Ossification.
NR2F2 regulates bone marrow-derived mesenchymal stem cell-promoted proliferation of Reh cells.
MIF Plays a Key Role in Regulating Tissue-Specific Chondro-Osteogenic Differentiation Fate of Human Cartilage Endplate Stem Cells under Hypoxia.
Adipose-derived Mesenchymal Stem Cells Are Phenotypically Superior for Regeneration in the Setting of Osteonecrosis of the Femoral Head.
Human adipose-derived stem cells partially rescue the stroke syndromes by promoting spatial learning and memory in mouse middle cerebral artery occlusion model.
Efficacy of autologous stem cell-based therapy for osteonecrosis of the femoral head in sickle cell disease: a five-year follow-up study.
Human Gingival Integration-Free iPSCs; a Source for MSC-Like Cells.
Impaired function of bone marrow stromal cells in systemic mastocytosis.
Modeling familial cancer with induced pluripotent stem cells.
Undifferentiated human adipose-derived stromal/stem cells loaded onto wet-spun starch-polycaprolactone scaffolds enhance bone regeneration: nude mice calvarial defect in vivo study.
Epithelial cell differentiation of human mesenchymal stromal cells in decellularized lung scaffolds.
Lipopolysaccharide activates Toll-like receptor 4 (TLR4)-mediated NF-κB signaling pathway and proinflammatory response in human pericytes.
Resolving cancer-stroma interfacial signalling and interventions with micropatterned tumour-stromal assays.
Hypoxia-cultured human adipose-derived mesenchymal stem cells are non-oncogenic and have enhanced viability, motility, and tropism to brain cancer.
Serum- and growth-factor-free three-dimensional culture system supports cartilage tissue formation by promoting collagen synthesis via Sox9-Col2a1 interaction.
Differentiation of human adipose-derived stem cells into neuron-like cells which are compatible with photocurable three-dimensional scaffolds.
Low-oxygen tension and IGF-I promote proliferation and multipotency of placental mesenchymal stem cells (PMSCs) from different gestations via distinct signaling pathways.
In vitro mesenchymal trilineage differentiation and extracellular matrix production by adipose and bone marrow derived adult equine multipotent stromal cells on a collagen scaffold.
Early intervention with gene-modified mesenchymal stem cells overexpressing interleukin-4 enhances anti-inflammatory responses and functional recovery in experimental autoimmune demyelination.
Tumor immunotherapy using gene-modified human mesenchymal stem cells loaded into synthetic extracellular matrix scaffolds.
Neuroprotective effects of mesenchymal stem cells derived from human embryonic stem cells in transient focal cerebral ischemia in rats.
Electrophysiological properties of human adipose tissue-derived stem cells.
Gao Y, Duan R, Li H, Jiang L, Tao T, Liu X, Zhu L, Li Z, Chen B, Zheng S, Lin X, Su W
iScience 2023 May 19;26(5):106729
iScience 2023 May 19;26(5):106729
Neuronal Cell Differentiation of Human Dental Pulp Stem Cells on Synthetic Polymeric Surfaces Coated With ECM Proteins.
Gao Y, Tian Z, Liu Q, Wang T, Ban LK, Lee HH, Umezawa A, Almansour AI, Arumugam N, Kumar RS, Ye Q, Higuchi A, Chen H, Sung TC
Frontiers in cell and developmental biology 2022;10:893241
Frontiers in cell and developmental biology 2022;10:893241
Mesenchymal stem cells derived from patients with premature aging syndromes display hallmarks of physiological aging.
Trani JP, Chevalier R, Caron L, El Yazidi C, Broucqsault N, Toury L, Thomas M, Annab K, Binetruy B, De Sandre-Giovannoli A, Levy N, Magdinier F, Robin JD
Life science alliance 2022 Sep 14;5(12)
Life science alliance 2022 Sep 14;5(12)
Carboxymethyl chitin or chitosan for osteoinduction effect on the human periodontal ligament stem cells.
Fan C, Li Z, Ji Q, Sun H, Liang Y, Yang P
Dental materials journal 2022 May 31;41(3):392-401
Dental materials journal 2022 May 31;41(3):392-401
Exosomes derived from stem cells of human deciduous exfoliated teeth inhibit angiogenesis in vivo and in vitro via the transfer of miR-100-5p and miR-1246.
Liu P, Zhang Q, Mi J, Wang S, Xu Q, Zhuang D, Chen W, Liu C, Zhang L, Guo J, Wu X
Stem cell research & therapy 2022 Mar 3;13(1):89
Stem cell research & therapy 2022 Mar 3;13(1):89
Mechanical Compression by Simulating Orthodontic Tooth Movement in an In Vitro Model Modulates Phosphorylation of AKT and MAPKs via TLR4 in Human Periodontal Ligament Cells.
Roth CE, Craveiro RB, Niederau C, Malyaran H, Neuss S, Jankowski J, Wolf M
International journal of molecular sciences 2022 Jul 22;23(15)
International journal of molecular sciences 2022 Jul 22;23(15)
In-cytoplasm mitochondrial transplantation for mesenchymal stem cells engineering and tissue regeneration.
Yao X, Ma Y, Zhou W, Liao Y, Jiang Z, Lin J, He Q, Wu H, Wei W, Wang X, Björklund M, Ouyang H
Bioengineering & translational medicine 2022 Jan;7(1):e10250
Bioengineering & translational medicine 2022 Jan;7(1):e10250
Induced Neurodifferentiation of hBM-MSCs through Activation of the ERK/CREB Pathway via Pulsed Electromagnetic Fields and Physical Stimulation Promotes Neurogenesis in Cerebral Ischemic Models.
Park HJ, Choi JH, Nam MH, Seo YK
International journal of molecular sciences 2022 Jan 21;23(3)
International journal of molecular sciences 2022 Jan 21;23(3)
Isolation and characterization mesenchymal stem cells from red panda (Ailurus fulgens styani) endometrium.
Wang DH, Wu XM, Chen JS, Cai ZG, An JH, Zhang MY, Li Y, Li FP, Hou R, Liu YL
Conservation physiology 2022 Jan 1;10(1):coac004
Conservation physiology 2022 Jan 1;10(1):coac004
Production enhancement of human adipose-derived mesenchymal stem cells by low-intensity ultrasound stimulation.
Min S, Byeon Y, Kim M, Lee Y, Lee SH, Lee Y, Farooqi HMU, Lee HK, Paeng DG
Scientific reports 2022 Dec 21;12(1):22041
Scientific reports 2022 Dec 21;12(1):22041
Aberrant Expression of COX-2 and FOXG1 in Infrapatellar Fat Pad-Derived ASCs from Pre-Diabetic Donors.
O'Donnell BT, Monjure TA, Al-Ghadban S, Ives CJ, L'Ecuyer MP, Rhee C, Romero-Lopez M, Li Z, Goodman SB, Lin H, Tuan RS, Bunnell BA
Cells 2022 Aug 1;11(15)
Cells 2022 Aug 1;11(15)
Human Amniotic Mesenchymal Stem Cells Inhibit aGVHD by Regulating Balance of Treg and T Effector Cells.
Gao Y, Li W, Bu X, Xu Y, Cai S, Zhong J, Du M, Sun H, Huang L, He Y, Hu X, Liu Q, Jin H, Wang Q, Ping B
Journal of inflammation research 2021;14:3985-3999
Journal of inflammation research 2021;14:3985-3999
Development, characterization, and hematopoietic differentiation of Griscelli syndrome type 2 induced pluripotent stem cells.
Güney-Esken G, Erol ÖD, Pervin B, Gürhan Sevinç G, Önder T, Bilgiç E, Korkusuz P, Günel-Özcan A, Uçkan-Çetinkaya D, Aerts-Kaya F
Stem cell research & therapy 2021 May 13;12(1):287
Stem cell research & therapy 2021 May 13;12(1):287
Therapeutic potential of human umbilical cord mesenchymal stem cells on aortic atherosclerotic plaque in a high-fat diet rabbit model.
Li Y, Shi G, Han Y, Shang H, Li H, Liang W, Zhao W, Bai L, Qin C
Stem cell research & therapy 2021 Jul 15;12(1):407
Stem cell research & therapy 2021 Jul 15;12(1):407
Glycemic control by umbilical cord-derived mesenchymal stem cells promotes effects of fasting-mimicking diet on type 2 diabetic mice.
Zhao N, Gao YF, Bao L, Lei J, An HX, Pu FX, Cheng RP, Chen J, Ni H, Sui BD, Ji FP, Hu CH
Stem cell research & therapy 2021 Jul 13;12(1):395
Stem cell research & therapy 2021 Jul 13;12(1):395
Differentiation potential and mRNA profiles of human dedifferentiated adipose cells and adipose‑derived stem cells from young donors.
Nie F, Bi H, Zhang C, Ding P
Molecular medicine reports 2021 Jan;23(1)
Molecular medicine reports 2021 Jan;23(1)
TRPA1 triggers hyperalgesia and inflammation after tooth bleaching.
Chen C, Huang X, Zhu W, Ding C, Huang P, Li R
Scientific reports 2021 Aug 31;11(1):17418
Scientific reports 2021 Aug 31;11(1):17418
The Secretome Derived From Mesenchymal Stromal Cells Cultured in a Xeno-Free Medium Promotes Human Cartilage Recovery in vitro.
Palamà MEF, Shaw GM, Carluccio S, Reverberi D, Sercia L, Persano L, Pisignano D, Cortese K, Barry FP, Murphy JM, Gentili C
Frontiers in bioengineering and biotechnology 2020;8:90
Frontiers in bioengineering and biotechnology 2020;8:90
Down-Regulated Exosomal MicroRNA-221 - 3p Derived From Senescent Mesenchymal Stem Cells Impairs Heart Repair.
Sun L, Zhu W, Zhao P, Zhang J, Lu Y, Zhu Y, Zhao W, Liu Y, Chen Q, Zhang F
Frontiers in cell and developmental biology 2020;8:263
Frontiers in cell and developmental biology 2020;8:263
Differentiation Potential of Early- and Late-Passage Adipose-Derived Mesenchymal Stem Cells Cultured under Hypoxia and Normoxia.
Zhao AG, Shah K, Freitag J, Cromer B, Sumer H
Stem cells international 2020;2020:8898221
Stem cells international 2020;2020:8898221
Vasculogenesis from Human Dental Pulp Stem Cells Grown in Matrigel with Fully Defined Serum-Free Culture Media.
Luzuriaga J, Irurzun J, Irastorza I, Unda F, Ibarretxe G, Pineda JR
Biomedicines 2020 Nov 9;8(11)
Biomedicines 2020 Nov 9;8(11)
Apoptotic bodies derived from mesenchymal stem cells promote cutaneous wound healing via regulating the functions of macrophages.
Liu J, Qiu X, Lv Y, Zheng C, Dong Y, Dou G, Zhu B, Liu A, Wang W, Zhou J, Liu S, Liu S, Gao B, Jin Y
Stem cell research & therapy 2020 Nov 27;11(1):507
Stem cell research & therapy 2020 Nov 27;11(1):507
The Effect of Angiotensin II, Retinoic Acid, EGCG, and Vitamin C on the Cardiomyogenic Differentiation Induction of Human Amniotic Fluid-Derived Mesenchymal Stem Cells.
Gasiūnienė M, Valatkaitė E, Navakauskaitė A, Navakauskienė R
International journal of molecular sciences 2020 Nov 19;21(22)
International journal of molecular sciences 2020 Nov 19;21(22)
Resveratrol rescues TNF‑α‑induced inhibition of osteogenesis in human periodontal ligament stem cells via the ERK1/2 pathway.
Yuan J, Wang X, Ma D, Gao H, Zheng D, Zhang J
Molecular medicine reports 2020 May;21(5):2085-2094
Molecular medicine reports 2020 May;21(5):2085-2094
Migration Inhibitory Factor in Conditioned Medium from Human Umbilical Cord Blood-Derived Mesenchymal Stromal Cells Stimulates Hair Growth.
Oh HA, Kwak J, Kim BJ, Jin HJ, Park WS, Choi SJ, Oh W, Um S
Cells 2020 May 28;9(6)
Cells 2020 May 28;9(6)
Chondrogenic Differentiation from Induced Pluripotent Stem Cells Using Non-Viral Minicircle Vectors.
Rim YA, Nam Y, Park N, Jung H, Lee K, Lee J, Ju JH
Cells 2020 Mar 1;9(3)
Cells 2020 Mar 1;9(3)
ZKSCAN3 counteracts cellular senescence by stabilizing heterochromatin.
Hu H, Ji Q, Song M, Ren J, Liu Z, Wang Z, Liu X, Yan K, Hu J, Jing Y, Wang S, Zhang W, Liu GH, Qu J
Nucleic acids research 2020 Jun 19;48(11):6001-6018
Nucleic acids research 2020 Jun 19;48(11):6001-6018
Rescue of premature aging defects in Cockayne syndrome stem cells by CRISPR/Cas9-mediated gene correction.
Wang S, Min Z, Ji Q, Geng L, Su Y, Liu Z, Hu H, Wang L, Zhang W, Suzuiki K, Huang Y, Zhang P, Tang TS, Qu J, Yu Y, Liu GH, Qiao J
Protein & cell 2020 Jan;11(1):1-22
Protein & cell 2020 Jan;11(1):1-22
Impairment of endothelial progenitor cells function in patient with mustard gas intoxication.
Siavashi V, Cheraghi H, Pourmohammad P, Nooshirvani P, Abdolahi S, Solghani A, Nassiri SM, Ghazanfari T
Inhalation toxicology 2020 Feb;32(3):131-140
Inhalation toxicology 2020 Feb;32(3):131-140
PF-127 hydrogel plus sodium ascorbyl phosphate improves Wharton's jelly mesenchymal stem cell-mediated skin wound healing in mice.
Deng Q, Huang S, Wen J, Jiao Y, Su X, Shi G, Huang J
Stem cell research & therapy 2020 Apr 3;11(1):143
Stem cell research & therapy 2020 Apr 3;11(1):143
Therapeutic Effects of Human Urine-Derived Stem Cells in a Rat Model of Cisplatin-Induced Acute Kidney Injury In Vivo and In Vitro.
Sun B, Luo X, Yang C, Liu P, Yang Y, Dong X, Yang Z, Xu J, Zhang Y, Li L
Stem cells international 2019;2019:8035076
Stem cells international 2019;2019:8035076
Investigating the potential of the secretome of mesenchymal stem cells derived from sickle cell disease patients.
Ribeiro TO, Silveira BM, Meira MC, Carreira ACO, Sogayar MC, Meyer R, Fortuna V
PloS one 2019;14(10):e0222093
PloS one 2019;14(10):e0222093
Ultrastructural morphology is distinct among primary progenitor cell isolates from normal, inflamed, and cryopreserved equine hoof tissue and CD105(+)K14(+) progenitor cells.
Yang Q, Lopez MJ
In vitro cellular & developmental biology. Animal 2019 Sep;55(8):641-655
In vitro cellular & developmental biology. Animal 2019 Sep;55(8):641-655
Telomere-dependent and telomere-independent roles of RAP1 in regulating human stem cell homeostasis.
Zhang X, Liu Z, Liu X, Wang S, Zhang Y, He X, Sun S, Ma S, Shyh-Chang N, Liu F, Wang Q, Wang X, Liu L, Zhang W, Song M, Liu GH, Qu J
Protein & cell 2019 Sep;10(9):649-667
Protein & cell 2019 Sep;10(9):649-667
TGF‑β induces periodontal ligament stem cell senescence through increase of ROS production.
Fan C, Ji Q, Zhang C, Xu S, Sun H, Li Z
Molecular medicine reports 2019 Oct;20(4):3123-3130
Molecular medicine reports 2019 Oct;20(4):3123-3130
High-yield isolation of menstrual blood-derived endometrial stem cells by direct red blood cell lysis treatment.
Sun Y, Ren Y, Yang F, He Y, Liang S, Guan L, Cheng F, Liu Y, Lin J
Biology open 2019 May 2;8(5)
Biology open 2019 May 2;8(5)
Better therapeutic potential of bone marrow-derived mesenchymal stem cells compared with chorionic villi-derived mesenchymal stem cells in airway injury model.
Ji S, Wu C, Tong L, Wang L, Zhou J, Chen C, Song Y
Regenerative medicine 2019 Mar;14(3):165-177
Regenerative medicine 2019 Mar;14(3):165-177
Stabilizing heterochromatin by DGCR8 alleviates senescence and osteoarthritis.
Deng L, Ren R, Liu Z, Song M, Li J, Wu Z, Ren X, Fu L, Li W, Zhang W, Guillen P, Izpisua Belmonte JC, Chan P, Qu J, Liu GH
Nature communications 2019 Jul 26;10(1):3329
Nature communications 2019 Jul 26;10(1):3329
Bone mesenchymal stromal cells exhibit functional inhibition but no chromosomal aberrations in chronic myelogenous leukemia.
Xie J, Chen J, Wang B, He X, Huang H
Oncology letters 2019 Jan;17(1):999-1007
Oncology letters 2019 Jan;17(1):999-1007
Visfatin Mediates Malignant Behaviors through Adipose-Derived Stem Cells Intermediary in Breast Cancer.
Huang JY, Wang YY, Lo S, Tseng LM, Chen DR, Wu YC, Hou MF, Yuan SF
Cancers 2019 Dec 20;12(1)
Cancers 2019 Dec 20;12(1)
Up-regulation of FOXD1 by YAP alleviates senescence and osteoarthritis.
Fu L, Hu Y, Song M, Liu Z, Zhang W, Yu FX, Wu J, Wang S, Izpisua Belmonte JC, Chan P, Qu J, Tang F, Liu GH
PLoS biology 2019 Apr;17(4):e3000201
PLoS biology 2019 Apr;17(4):e3000201
ATF6 safeguards organelle homeostasis and cellular aging in human mesenchymal stem cells.
Wang S, Hu B, Ding Z, Dang Y, Wu J, Li D, Liu X, Xiao B, Zhang W, Ren R, Lei J, Hu H, Chen C, Chan P, Li D, Qu J, Tang F, Liu GH
Cell discovery 2018;4:2
Cell discovery 2018;4:2
The ICAM-1 expression level determines the susceptibility of human endothelial cells to simulated microgravity.
Buravkova LB, Rudimov EG, Andreeva ER, Grigoriev AI
Journal of cellular biochemistry 2018 Mar;119(3):2875-2885
Journal of cellular biochemistry 2018 Mar;119(3):2875-2885
BNIP3/Bcl-2-mediated apoptosis induced by cyclic tensile stretch in human cartilage endplate-derived stem cells.
Yuan C, Pu L, He Z, Wang J
Experimental and therapeutic medicine 2018 Jan;15(1):235-241
Experimental and therapeutic medicine 2018 Jan;15(1):235-241
Isolation and characterization of stem cells from differentially degenerated human lumbar zygapophyseal articular cartilage.
Xiao L, Xu S, Wang X, Jin Z, Wang J, Yang B, Xu H
Molecular medicine reports 2018 Dec;18(6):5751-5759
Molecular medicine reports 2018 Dec;18(6):5751-5759
Human umbilical cord-derived mesenchymal stem cells ameliorate the enteropathy of food allergies in mice.
Yan N, Xu J, Zhao C, Wu Y, Gao F, Li C, Zhou W, Xiao T, Zhou X, Shao Q, Xia S
Experimental and therapeutic medicine 2018 Dec;16(6):4445-4456
Experimental and therapeutic medicine 2018 Dec;16(6):4445-4456
Antagonizing CD105 enhances radiation sensitivity in prostate cancer.
Madhav A, Andres A, Duong F, Mishra R, Haldar S, Liu Z, Angara B, Gottlieb R, Zumsteg ZS, Bhowmick NA
Oncogene 2018 Aug;37(32):4385-4397
Oncogene 2018 Aug;37(32):4385-4397
Differential stem cell aging kinetics in Hutchinson-Gilford progeria syndrome and Werner syndrome.
Wu Z, Zhang W, Song M, Wang W, Wei G, Li W, Lei J, Huang Y, Sang Y, Chan P, Chen C, Qu J, Suzuki K, Belmonte JCI, Liu GH
Protein & cell 2018 Apr;9(4):333-350
Protein & cell 2018 Apr;9(4):333-350
Effects of acute exposure to low-dose radiation on the characteristics of human bone marrow mesenchymal stromal/stem cells.
Fujishiro A, Miura Y, Iwasa M, Fujii S, Sugino N, Andoh A, Hirai H, Maekawa T, Ichinohe T
Inflammation and regeneration 2017;37:19
Inflammation and regeneration 2017;37:19
Insulin-Like Growth Factor Binding Protein-6 Alters Skeletal Muscle Differentiation of Human Mesenchymal Stem Cells.
Aboalola D, Han VKM
Stem cells international 2017;2017:2348485
Stem cells international 2017;2017:2348485
Endoglin: a novel target for therapeutic intervention in acute leukemias revealed in xenograft mouse models.
Dourado KMC, Baik J, Oliveira VKP, Beltrame M, Yamamoto A, Theuer CP, Figueiredo CAV, Verneris MR, Perlingeiro RCR
Blood 2017 May 4;129(18):2526-2536
Blood 2017 May 4;129(18):2526-2536
A high-yield isolation and enrichment strategy for human lung microvascular endothelial cells.
Gaskill C, Majka SM
Pulmonary circulation 2017 Mar;7(1):108-116
Pulmonary circulation 2017 Mar;7(1):108-116
The Improvement of Respiratory Performance After Phototherapy-Induced EPC Mobilization in Preterm Infants With RDS.
Siavashi V, Asadian S, Taheri-Asl M, Babaei H, Keshavarz S, Bazaei M, Nassiri SM
Journal of cellular biochemistry 2017 Mar;118(3):594-604
Journal of cellular biochemistry 2017 Mar;118(3):594-604
Human umbilical cord mesenchymal stem cells improve the reserve function of perimenopausal ovary via a paracrine mechanism.
Li J, Mao Q, He J, She H, Zhang Z, Yin C
Stem cell research & therapy 2017 Mar 9;8(1):55
Stem cell research & therapy 2017 Mar 9;8(1):55
Elimination of undifferentiated human embryonic stem cells by cardiac glycosides.
Lin YT, Wang CK, Yang SC, Hsu SC, Lin H, Chang FP, Kuo TC, Shen CN, Chiang PM, Hsiao M, Lu FL, Lu J
Scientific reports 2017 Jul 13;7(1):5289
Scientific reports 2017 Jul 13;7(1):5289
Circulation Enrichment of Functional Endothelial Progenitor Cells by Infantile Phototherapy.
Siavashi V, Asadian S, Sharifi A, Esmaeilivand M, Norouzinia R, Azadbakht M, Nassiri SM
Journal of cellular biochemistry 2017 Feb;118(2):330-340
Journal of cellular biochemistry 2017 Feb;118(2):330-340
Feasibility of placenta-derived mesenchymal stem cells as a tool for studying pregnancy-related disorders.
Fuchi N, Miura K, Doi H, Li TS, Masuzaki H
Scientific reports 2017 Apr 12;7:46220
Scientific reports 2017 Apr 12;7:46220
A Member of the Nuclear Receptor Superfamily, Designated as NR2F2, Supports the Self-Renewal Capacity and Pluripotency of Human Bone Marrow-Derived Mesenchymal Stem Cells.
Zhu N, Wang H, Wang B, Wei J, Shan W, Feng J, Huang H
Stem cells international 2016;2016:5687589
Stem cells international 2016;2016:5687589
CRISPR/Cas9-based genetic correction for recessive dystrophic epidermolysis bullosa.
Webber BR, Osborn MJ, McElroy AN, Twaroski K, Lonetree CL, DeFeo AP, Xia L, Eide C, Lees CJ, McElmurry RT, Riddle MJ, Kim CJ, Patel DD, Blazar BR, Tolar J
NPJ Regenerative medicine 2016;1:16014-
NPJ Regenerative medicine 2016;1:16014-
Influence of aging on the quantity and quality of human cardiac stem cells.
Nakamura T, Hosoyama T, Kawamura D, Takeuchi Y, Tanaka Y, Samura M, Ueno K, Nishimoto A, Kurazumi H, Suzuki R, Ito H, Sakata K, Mikamo A, Li TS, Hamano K
Scientific reports 2016 Mar 7;6:22781
Scientific reports 2016 Mar 7;6:22781
Angiogenic activity mediates bone repair from human pluripotent stem cell-derived osteogenic cells.
Zou L, Chen Q, Quanbeck Z, Bechtold JE, Kaufman DS
Scientific reports 2016 Mar 16;6:22868
Scientific reports 2016 Mar 16;6:22868
Potency of umbilical cord blood- and Wharton's jelly-derived mesenchymal stem cells for scarless wound healing.
Doi H, Kitajima Y, Luo L, Yan C, Tateishi S, Ono Y, Urata Y, Goto S, Mori R, Masuzaki H, Shimokawa I, Hirano A, Li TS
Scientific reports 2016 Jan 5;6:18844
Scientific reports 2016 Jan 5;6:18844
SIRT6 safeguards human mesenchymal stem cells from oxidative stress by coactivating NRF2.
Pan H, Guan D, Liu X, Li J, Wang L, Wu J, Zhou J, Zhang W, Ren R, Zhang W, Li Y, Yang J, Hao Y, Yuan T, Yuan G, Wang H, Ju Z, Mao Z, Li J, Qu J, Tang F, Liu GH
Cell research 2016 Feb;26(2):190-205
Cell research 2016 Feb;26(2):190-205
Lymphatic Contribution to the Cellular Niche in Heterotopic Ossification.
Loder S, Agarwal S, Sorkin M, Breuler C, Li J, Peterson J, Gardenier J, Hsieh HH, Wang SC, Mehrara BJ, Levi B
Annals of surgery 2016 Dec;264(6):1174-1180
Annals of surgery 2016 Dec;264(6):1174-1180
NR2F2 regulates bone marrow-derived mesenchymal stem cell-promoted proliferation of Reh cells.
Zhu N, Wang H, Wei J, Wang B, Shan W, Lai X, Zhao Y, Yu J, Huang H
Molecular medicine reports 2016 Aug;14(2):1351-6
Molecular medicine reports 2016 Aug;14(2):1351-6
MIF Plays a Key Role in Regulating Tissue-Specific Chondro-Osteogenic Differentiation Fate of Human Cartilage Endplate Stem Cells under Hypoxia.
Yao Y, Deng Q, Song W, Zhang H, Li Y, Yang Y, Fan X, Liu M, Shang J, Sun C, Tang Y, Jin X, Liu H, Huang B, Zhou Y
Stem cell reports 2016 Aug 9;7(2):249-62
Stem cell reports 2016 Aug 9;7(2):249-62
Adipose-derived Mesenchymal Stem Cells Are Phenotypically Superior for Regeneration in the Setting of Osteonecrosis of the Femoral Head.
Wyles CC, Houdek MT, Crespo-Diaz RJ, Norambuena GA, Stalboerger PG, Terzic A, Behfar A, Sierra RJ
Clinical orthopaedics and related research 2015 Oct;473(10):3080-90
Clinical orthopaedics and related research 2015 Oct;473(10):3080-90
Human adipose-derived stem cells partially rescue the stroke syndromes by promoting spatial learning and memory in mouse middle cerebral artery occlusion model.
Zhou F, Gao S, Wang L, Sun C, Chen L, Yuan P, Zhao H, Yi Y, Qin Y, Dong Z, Cao L, Ren H, Zhu L, Li Q, Lu B, Liang A, Xu GT, Zhu H, Gao Z, Ma J, Xu J, Chen X
Stem cell research & therapy 2015 May 9;6(1):92
Stem cell research & therapy 2015 May 9;6(1):92
Efficacy of autologous stem cell-based therapy for osteonecrosis of the femoral head in sickle cell disease: a five-year follow-up study.
Daltro GC, Fortuna V, de Souza ES, Salles MM, Carreira AC, Meyer R, Freire SM, Borojevic R
Stem cell research & therapy 2015 May 29;6(1):110
Stem cell research & therapy 2015 May 29;6(1):110
Human Gingival Integration-Free iPSCs; a Source for MSC-Like Cells.
Umezaki Y, Hashimoto Y, Nishishita N, Kawamata S, Baba S
International journal of molecular sciences 2015 Jun 15;16(6):13633-48
International journal of molecular sciences 2015 Jun 15;16(6):13633-48
Impaired function of bone marrow stromal cells in systemic mastocytosis.
Nemeth K, Wilson TM, Ren JJ, Sabatino M, Stroncek DM, Krepuska M, Bai Y, Robey PG, Metcalfe DD, Mezey E
Stem cell research 2015 Jul;15(1):42-53
Stem cell research 2015 Jul;15(1):42-53
Modeling familial cancer with induced pluripotent stem cells.
Lee DF, Su J, Kim HS, Chang B, Papatsenko D, Zhao R, Yuan Y, Gingold J, Xia W, Darr H, Mirzayans R, Hung MC, Schaniel C, Lemischka IR
Cell 2015 Apr 9;161(2):240-54
Cell 2015 Apr 9;161(2):240-54
Undifferentiated human adipose-derived stromal/stem cells loaded onto wet-spun starch-polycaprolactone scaffolds enhance bone regeneration: nude mice calvarial defect in vivo study.
Carvalho PP, Leonor IB, Smith BJ, Dias IR, Reis RL, Gimble JM, Gomes ME
Journal of biomedical materials research. Part A 2014 Sep;102(9):3102-11
Journal of biomedical materials research. Part A 2014 Sep;102(9):3102-11
Epithelial cell differentiation of human mesenchymal stromal cells in decellularized lung scaffolds.
Mendez JJ, Ghaedi M, Steinbacher D, Niklason LE
Tissue engineering. Part A 2014 Jun;20(11-12):1735-46
Tissue engineering. Part A 2014 Jun;20(11-12):1735-46
Lipopolysaccharide activates Toll-like receptor 4 (TLR4)-mediated NF-κB signaling pathway and proinflammatory response in human pericytes.
Guijarro-Muñoz I, Compte M, Álvarez-Cienfuegos A, Álvarez-Vallina L, Sanz L
The Journal of biological chemistry 2014 Jan 24;289(4):2457-68
The Journal of biological chemistry 2014 Jan 24;289(4):2457-68
Resolving cancer-stroma interfacial signalling and interventions with micropatterned tumour-stromal assays.
Shen K, Luk S, Hicks DF, Elman JS, Bohr S, Iwamoto Y, Murray R, Pena K, Wang F, Seker E, Weissleder R, Yarmush ML, Toner M, Sgroi D, Parekkadan B
Nature communications 2014 Dec 9;5:5662
Nature communications 2014 Dec 9;5:5662
Hypoxia-cultured human adipose-derived mesenchymal stem cells are non-oncogenic and have enhanced viability, motility, and tropism to brain cancer.
Feng Y, Zhu M, Dangelmajer S, Lee YM, Wijesekera O, Castellanos CX, Denduluri A, Chaichana KL, Li Q, Zhang H, Levchenko A, Guerrero-Cazares H, Quiñones-Hinojosa A
Cell death & disease 2014 Dec 11;5(12):e1567
Cell death & disease 2014 Dec 11;5(12):e1567
Serum- and growth-factor-free three-dimensional culture system supports cartilage tissue formation by promoting collagen synthesis via Sox9-Col2a1 interaction.
Ahmed N, Iu J, Brown CE, Taylor DW, Kandel RA
Tissue engineering. Part A 2014 Aug;20(15-16):2224-33
Tissue engineering. Part A 2014 Aug;20(15-16):2224-33
Differentiation of human adipose-derived stem cells into neuron-like cells which are compatible with photocurable three-dimensional scaffolds.
Gao S, Zhao P, Lin C, Sun Y, Wang Y, Zhou Z, Yang D, Wang X, Xu H, Zhou F, Cao L, Zhou W, Ning K, Chen X, Xu J
Tissue engineering. Part A 2014 Apr;20(7-8):1271-84
Tissue engineering. Part A 2014 Apr;20(7-8):1271-84
Low-oxygen tension and IGF-I promote proliferation and multipotency of placental mesenchymal stem cells (PMSCs) from different gestations via distinct signaling pathways.
Youssef A, Iosef C, Han VK
Endocrinology 2014 Apr;155(4):1386-97
Endocrinology 2014 Apr;155(4):1386-97
In vitro mesenchymal trilineage differentiation and extracellular matrix production by adipose and bone marrow derived adult equine multipotent stromal cells on a collagen scaffold.
Xie L, Zhang N, Marsano A, Vunjak-Novakovic G, Zhang Y, Lopez MJ
Stem cell reviews and reports 2013 Dec;9(6):858-72
Stem cell reviews and reports 2013 Dec;9(6):858-72
Early intervention with gene-modified mesenchymal stem cells overexpressing interleukin-4 enhances anti-inflammatory responses and functional recovery in experimental autoimmune demyelination.
Payne NL, Dantanarayana A, Sun G, Moussa L, Caine S, McDonald C, Herszfeld D, Bernard CC, Siatskas C
Cell adhesion & migration 2012 May-Jun;6(3):179-89
Cell adhesion & migration 2012 May-Jun;6(3):179-89
Tumor immunotherapy using gene-modified human mesenchymal stem cells loaded into synthetic extracellular matrix scaffolds.
Compte M, Cuesta AM, Sánchez-Martín D, Alonso-Camino V, Vicario JL, Sanz L, Alvarez-Vallina L
Stem cells (Dayton, Ohio) 2009 Mar;27(3):753-60
Stem cells (Dayton, Ohio) 2009 Mar;27(3):753-60
Neuroprotective effects of mesenchymal stem cells derived from human embryonic stem cells in transient focal cerebral ischemia in rats.
Liu YP, Seçkin H, Izci Y, Du ZW, Yan YP, Başkaya MK
Journal of cerebral blood flow and metabolism : official journal of the International Society of Cerebral Blood Flow and Metabolism 2009 Apr;29(4):780-91
Journal of cerebral blood flow and metabolism : official journal of the International Society of Cerebral Blood Flow and Metabolism 2009 Apr;29(4):780-91
Electrophysiological properties of human adipose tissue-derived stem cells.
Bai X, Ma J, Pan Z, Song YH, Freyberg S, Yan Y, Vykoukal D, Alt E
American journal of physiology. Cell physiology 2007 Nov;293(5):C1539-50
American journal of physiology. Cell physiology 2007 Nov;293(5):C1539-50
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Staining of U937 cells with Mouse IgG1 K Isotype Control PE (Product # 12-4714-81) (blue histogram) or Anti-Human CD105 (Endoglin) PE (purple histogram). Total viable cells, as determined by Fixable Viability Dye eFluor® 780, were used for analysis.
- Conjugate
- Yellow dye

Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 2 The phenotype of human CDCs varies according to the individual patient. The percentage of CD90- ( a ) and CD105-positive ( b ) cells was measured by flow cytometry. The average of CD90 and CD105 positivity was compared between CDCs from younger (=65 yrs) patients.
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 3 Characterization of MSC-like cells generated from iPSCs. ( A ) Morphology of HGF-iPSCs 1-1 at passage 45 cultured on GFRM with mTeSR1 ( left ). Morphology of MSC-like cells from HGF-iPSCs 1-1 differentiated (MSLCs 1-1) at passage 4 ( right ); ( B ) Flow cytometry analysis of MSC-related surface markers (CD44, CD73, CD90, and CD105), hematopoietic markers (CD34 and CD45), and pluripotent markers (SSEA3 and TRA1-60) on MSLCs 1-1 at passage 10. BMMSCs were used (passage 5) as positive control and HGF-iPSCs 1-1 (passage 45) as negative control; ( C ) MSC-like cells from HGF-iPSCs 1-2 (MSLCs 1-2) were differentiated at passage 23. Growth curve of BMMSCs, MSLCs 1-1 and 1-2. Scale bar = 400 mum. GFRM, Growth factor-reduced Matrigel.
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Fig. 1 Flow cytometry analysis of phenotype characterization of hUCMSCs. Phenotype of CD73, CD90, CD105, CD14, CD34, CD45, CD79a and HLA-DR of hUCMSCs was detected by flow cytometry. Intensity >= 95% represented strong expression while
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 1 Experimental diagram (A) , hADSC surface antigen profiling with FACS showed that hADSCs positively express CD29, CD44, CD73 and CD105, negatively express CD34 and CD45 (B) . FACS, flow activated cell sorting; hADSCs, human adipose-derived stem cells.
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
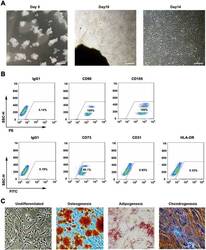
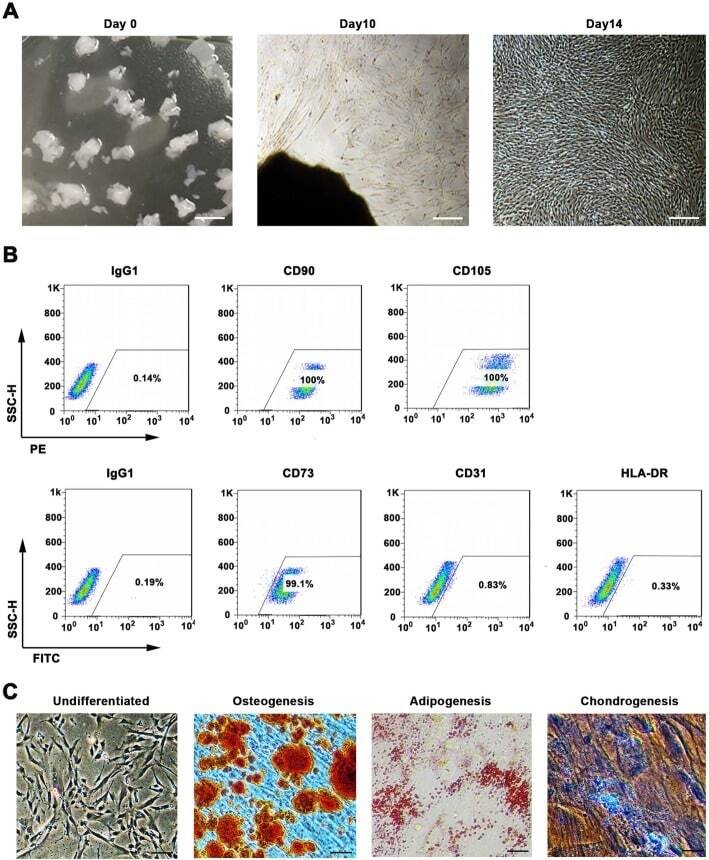
- Experimental details
- Fig. 1 WJMSCs isolation and characterization. a Primary cell isolation procedure from Wharton''s jelly tissue. The migrated cells exhibited typical fibroblast-like morphology. Scale bar, 500 mum. b Flow cytometry analysis of P4 cells using mesenchymal stem cell markers (CD90, CD105, CD73), endothelial cell marker (CD31), and MHC class II protein HLA-DR. Isotypic antibodies (IgG1-PE and IgG1-FITC) were used as negative controls. c Representative stained images show that the fourth passage WJMSCs could differentiate into osteocytes (Alizarin Red S), adipocytes (Oil Red O), and chondrocytes (Alcian blue). Scale bar, 100 mum
- Conjugate
- Yellow dye
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
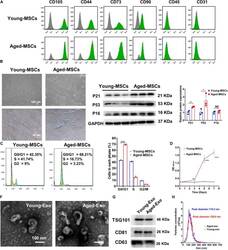
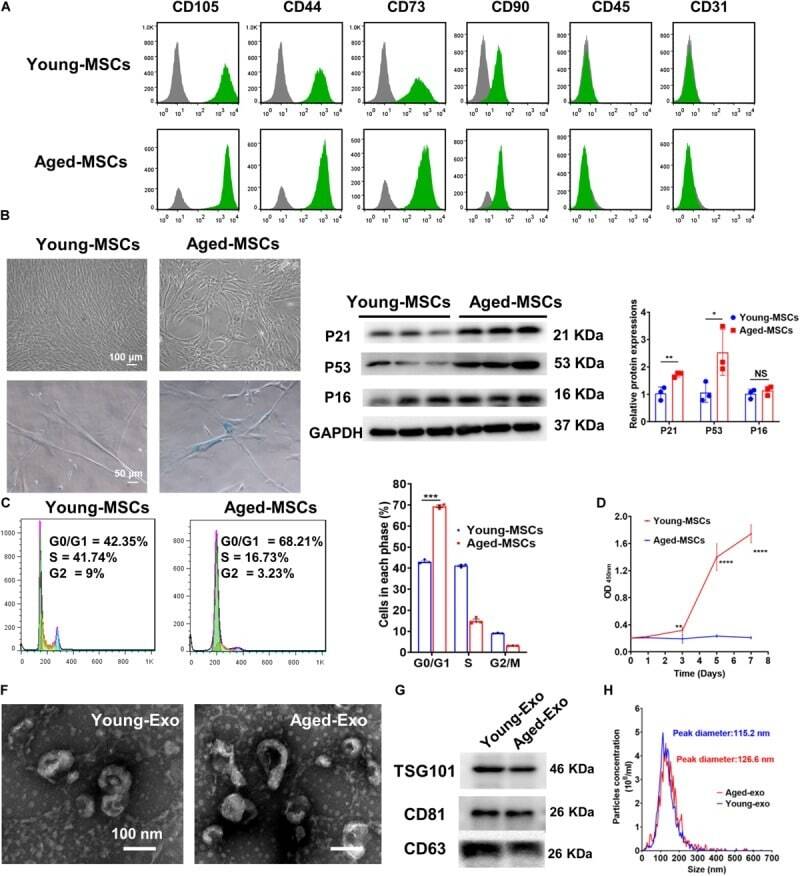
- Experimental details
- FIGURE 1 Characterization of young and aged MSCs and exosomes. (A) Surface marker profiling of young-MSCs and aged-MSCs. (B) SA-beta-Gal staining showed that senescence increased significantly in aged MSCs. (C) Representative immunoblot images and quantitative analysis of p21, p53, and p16 protein level in young and aged-MSCs. ( n = 3). (D) Quantitation of cell cycle phases by propidium iodide staining. ( n = 3). (E) The CCK-8 assay showed that aged MSCs grew more slowly than young MSCs. ( n = 6). (F) Young and aged exosomes were observed using TEM. (G) The exosome surface markers were analyzed by Western blot. (H) Nanoparticle tracking analysis was used to analyze the particle size and concentration of Young-Exo and Aged-Exo. * p < 0.05; ** p < 0.01; *** p < 0.001; **** p < 0.0001; NS, not significant.
- Conjugate
- Yellow dye
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Fig. 1 Characterization of a healthy donor and GS-2 BM-MSCs. a Morphology before and after differentiation of healthy donor (left, n =3) and GS-2 (right, n =3) BM-MSCs. MSCs were cultured in control medium (left lines) or adipogenic and osteogenic differentiation media for 21 days and were stained with Oil Red O (midline, ORO) for adipogenic differentiation and Alizarin Red S (right lines, ARS) for osteogenic differentiation. b Immunophenotype of a healthy donor and GS-2 BM-MSCs. Dot plots from representative BM-MSC samples of passage 3 GS-2 (upper lane) and healthy control (lower lane) BM-MSCs
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 2 Chondrogenesis using minicircle-transfected hiPSC-derived OG cells. ( a ) Scheme of chondrogenic differentiation process from hiPSCs. Minicircles were transfected after OG cells were induced. ( b ) Morphology of the hiPSC colony. ( c ) Morphology of the generated EBs. ( d ) Image of outgrowth cells derived from EBs attached to a gelatin-coated culture dish. ( e ) Morphology of OG cells before transfection. ( f ) Alizarin red-stained osteogenic cells differentiated from OG cells. ( g ) Oil red O staining image of adipogenic cells differentiated from OG cells. ( h ) Chondrogenic pellet generated from OG cells stained with alcian blue. Relative gene expression of ( i ) CD44, ( j ) CD73, ( k ) CD90, and ( l ) CD105 in OG cells. Percentage of ( m ) CD44, ( n ) CD73, ( o ) CD90, and ( p ) CD105 positive cells. ( q ) Fluorescence microscopy of mcMock-transfected OG cells. ( r ) Fluorescence microscopy of mcBMP2-transfected OG cells. ( s ) Fluorescence microscopy of mcTGFbeta3-transfected OG cells. ( t ) Percentage of OG cells transfected with each minicircle vectors. ( u ) Gel image of the PCR results against the insert of mcBMP2 and mcTGFbeta3 in transfected OG cells. Data are presented as mean +- standard deviation from three independent sets of experiments. Scale bars represents 200 mum. ** p < 0.01 and *** p < 0.001 indicate statistical significance. EB: embryonic body; OG: outgrowth; CDM: chondrogenic differentiation media; RFP: red fluorescence protein; MSC: mesenchym
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 1 SIRT6 -deficient hMSCs exhibit accelerated cell attrition. (A) Schematic representation of deletion of SIRT6 by removing exon 1 of SIRT6 gene via TALEN-based gene targeting technique. The donor vector contains a neomycin-resistant cassette (neo) allowing for positive selection, and the neo cassette was then removed from the SIRT6 gene locus. (B) Left panel: western blotting analysis of SIRT6 protein in hESCs. Protein extracts from wild-type (WT, SIRT6 +/+ ) and SIRT6 -deficient ( SIRT6 -/- ) hESCs were analyzed by western blotting using an anti-SIRT6 antibody. beta-actin was used as the loading control. Right panel: RT-PCR analysis of SIRT6 mRNA in hESCs. A pair of PCR primers spanning the junction region of SIRT6 mRNA exon 1 and exon 2 was used. 18S rRNA was used as the loading control. (C) Bright-field and SIRT6 immunofluorescence micrographs of WT and SIRT6 -deficient hESCs. DNA was stained by Hoechst 33342. Bright-field scale bar, 200 mum; immunofluorescence scale bar, 20 mum; zoom-field immunofluorescence scale bar, 10 mum. (D) Bright-field micrographs and FACS analysis of the surface markers CD105, CD73, and CD90 in WT and SIRT6 -deficient hMSCs. Scale bar, 100 mum. (E) Western blotting analysis of SIRT6 protein in hMSCs. Protein extracts from WT and SIRT6 -deficient hMSCs were analyzed by western blotting using anti-SIRT6 antibody. beta-actin was used as the loading control. (F) Immunofluorescence analysis showing the absence of SIRT6 protein in the nuclei of
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 1 Differentiation of hESCs and hiPSCs in osteogenic conditions. ( a ) Flow cytometric analysis of hESCs differentiated in various conditions, with osteogenic supplements (dexamethasone, ascorbic acid, and beta-glycerophosphate) in either 10% KOSR or 10% FBS, cultured on either gelatin, Matrigel, fibronectin, or by co-culture with MEFs or M210 stromal cells after 12 days of differentiation. A representative result is shown here from 3 independent experiments. ( b ) Summary of flow cytometric analysis of cell surface markers, CD73 and CD105, and intracellular osteogenic marker, osteocalcin, in the differentiated hESCs, UCBiPSCs and PBiPSCs cultured in the osteogenic medium with 10% FBS on gelatin at different passages. n = 3, *p < 0.05, One-way ANOVA with Tukey post-hoc tests for each comparing at each passage. ( c ) qRT-PCR analysis of osteogenic genes, RUNX2 , Osteocalcin ( OCN ) in the differentiated cells. The gene expression level was normalized by undifferentiated cells for each cell line, and data was summarized from 3 independent experiments, *p < 0.05, One-way ANOVA with Tukey post-hoc tests for each comparing at each passage.
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 2 Osteogenic phenotypes of osteogenic differentiated hESCs and hiPSCs in comparison with BM-MSC derived osteoprogenitor cells. ( a ) Summary of flow cytometric analysis of CD44, CD73, CD105 and CD146 positive populations in osteogenic differentiated cells at passage 6. ( b ) qRT-PCR analysis of three osteogenic genes in the differentiated cells at passage 6. Gene expression level was normalized by the expression level in BM-MSC-OS group. ( c ) ALP activity assay of the osteogenic differentiated cells (2 x 10 5 ) at passage 6 on day 3 after seeding. ( d ) Von Kossa staining and Alizarin Red staining of mineral deposition in the ECM around the differentiated cells (scale bar: 200 mum).
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
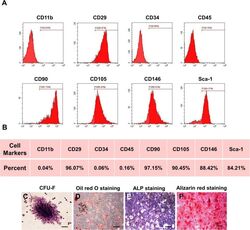
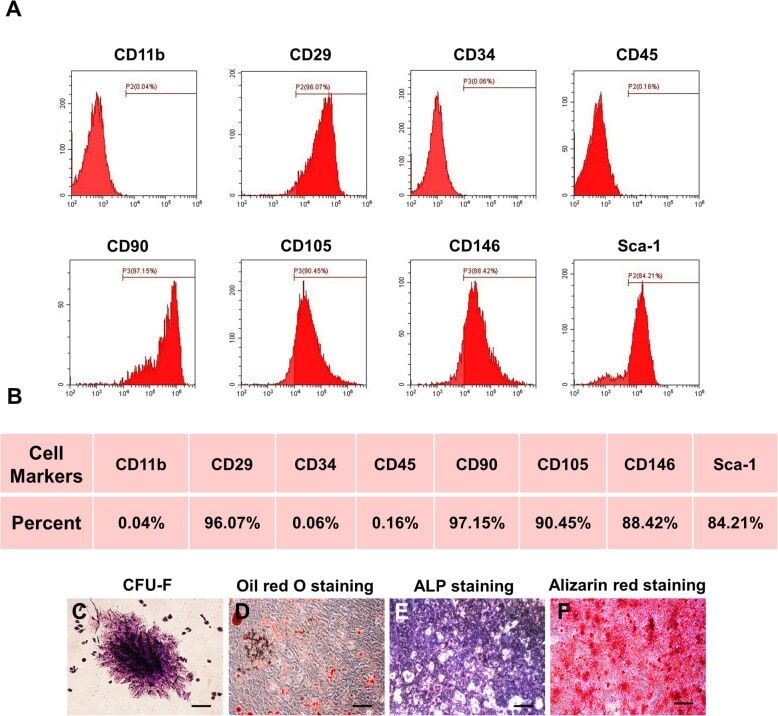
- Experimental details
- Fig. 1 Characterization of murine BMMSCs. a , b Flow cytometric analysis of the surface markers showed positive expression of CD29, CD90, CD105, CD146, and Sca-1, and negative expression of hematopoietic markers CD11b, CD34, and CD45. c Representative image of colony formation of murine BMMSCs. d Representative image of Oil Red O staining of murine BMMSCs. e Representative image of ALP staining of murine BMMSCs. f Representative image of Alizarin red staining of murine BMMSCs. Scale bar, 500 mum
- Conjugate
- Yellow dye
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 1 Identification of dental pulp stem cells (DPSCs). Human DPSCs were positive for the cell surface antigens CD73, CD90, and CD105, as well as negative for CD14, CD20, CD34, and CD45 demonstrated by flow cytometry ( A ). DPSCs were cultured under osteogenic ( B , 14 days) or adipogenic ( C , 21 days) conditions, and showed mineralized nodules and lipid clusters as revealed by alizarin red and oil red staining, respectively. Scale bar = 400 ( B ) or 100 ( C ) mum.
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 1 Identification of ADSCs. ( A ) ADSCs were isolated from the adipose tissue of breast tumors. After two to three passages, the expressions of ADSCs markers (CD90FITC, CD105PE, and CD44FITC) and the lack of CD34PE and CD45FITC were confirmed by flow cytometry. ( B ) The differentiation ability of ADSCs was tested by adipogenesis, osteogenesis, and chondrogenesis.
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Fig. 1 Characterization of UCSCs. A UCSCs display a spindle shaped and fibroblast-like morphology. B High UCSCs expression of CD90, CD29, CD73, and CD105, and low expression of HLA using flow cytometry
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 1 Characterization of human dental pulp stem cells (hDPSCs) cultured in Neurocult(tm) proliferation medium by flow cytometry. ( A ) Data quantification as representative histograms at 3 days in vitro (DIV) (gray) and 7DIV (black) for CD90, CD105 and CD73 mesenchymal stromal cell (MSC) markers, CD45 hematopoietic and CD31 endothelial markers ( n = 3). Data are represented as the average percentage and standard error of the mean (SEM+-), with respect to the total cell population. ( B ) Representative cytometry of CD90, CD105 and CD73 mesenchymal, CD45 hematopoietic and CD31 endothelial markers in hDPSCs cultured in Neurocult proliferation medium at 3DIV (gray filling), with respect to negative controls (no filling). ( C ) Representative cytometry of CD90, CD105 and CD73 mesenchymal, CD45 hematopoietic and CD31 endothelial markers in hDPSCs cultured in Neurocult proliferation medium at 7DIV (gray filling), with respect to negative controls (no filling). *: p < 0.05. Kruskal-Wallis with Dunn's post hoc test.
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Cell characteristics. Flow cytometric analysis of cEPCs for CD11b, CD14, CD45, CD105, CD115, and CD117 (A). Red line, isotype control; blue line, CD marker. Outgrowth of colony forming unit of endothelial cells (CFU-EC) after seeding of cEPCs on the methylcellulose medium containing VEGF and GM-CSF. The colony consisted of spindle-shaped cells with sprouting from the cluster center (B). Number of CFU-EC developed by cEPCs on the methylcellulose medium in each group (C). cEPCs did not give rise to CFU-GM.
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Cell characteristics. Flow cytometric analysis of cEPCs expanded on the fibronectin matrix for CD11b, CD14, CD45, CD105, and CD117 (A). Red line, isotype control; blue line, CD marker. Outgrowth of colony forming unit of endothelial cells (CFU-EC) after seeding of cEPCs on the methylcellulose medium containing VEGF and GM-CSF (B). Number of CFU-EC developed by cEPCs on the methylcellulose medium (C) (n = 6).
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 3 Acquisition and characterization of HGPS-MSCs and WS-MSCs . (A) FACS analysis of MSC-specific markers (CD73, CD90, CD105) in WT, heterozygous ( LMNA G608G/+ ), homozygous ( LMNA G608G/G608G ) and WRN -/- MSCs. (B) Left: characterization of adipogenesis potential of MSCs by Oil Red O staining. Right: Oil Red O positive areas were calculated by Image J. Data were presented as mean +- SEM, n = 3. * P < 0.05; ns, not significant. Scale bar, 100 mum. (C) Left: characterization of osteogenesis potential of MSCs by Von Kossa staining. Right: Von Kossa positive areas were calculated by Image J. Data were presented as mean +- SEM, n = 3. * P < 0.05; ns, not significant. Scale bar, 100 mum. (D) Left: characterization of chondrogenesis potential of MSCs by Toluidine Blue O staining. Right: the diameters of chondrocyte spheres were measured. Data were presented as mean +- SEM, n = 11. ns, not significant. Scale bar, 100 mum
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Fig. 1 Radiation-induced CD105 expression in prostate cancer cells supports radio-resistance. a Cell surface CD105 expression was measured in cell lines at 72 h following a dose range of irradiation (0, 2, 4, or 6 Gy). b The durability of cell surface CD105 expression in 22Rv1 was determined 0, 0.5, 4, 8, 24, 48, 72, 120, and 168 h following 4 Gy irradiation. CD105 cell surface expression fold change was normalized to levels expressed prior to irradiation. c The mRNA expression of CD105 ligands was measured at 0, 4, and 8 h post radiation by rtPCR. Expression was normalized to GAPDH and to the 0 h time point. d Western blot for phosphorylated Smad1/5 was measured in 22Rv1 cells in the presence or absence of serum starvation and treatment with 50 ng/ml BMP4 or TRC105. beta-actin expression served as the loading control. Molecular weight (kDa) is indicated. e Annexin-V expression was measured in 22Rv1 cells by FACS analysis 5 days following 4 Gy irradiation and treatment of IgG or TRC105. f Clonogenic assay was measured 10 days following irradiation of 22Rv1 and C4-2B cells in a dose range of 0-6 Gy in the presence of IgG or TRC105. Data are reported as a mean +- S.D. of three independent experiments (* p < 0.05, ** p < 0.01, *** p < 0.001 compared to control, unless otherwise indicated)
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 7 Safety analysis of gene-corrected CS-MSCs obtained under a cGMP-compliant condition . (A) FACS analysis indicated the expression of the cell surface markers CD73, CD90 and CD105 in CS-MSCs and GC-MSCs. (B) RT-qPCR analysis of the expression of pluripotency markers OCT4 , NANOG , and SOX2 in CS-MSCs and GC-MSCs. GC-iPSCs and CS-fibroblasts were used as positive and negative controls, respectively. Data are presented as the mean +- SEM, n = 3. (C) Immunostaining of the pluripotency marker NANOG in CS-MSCs and GC-MSCs. GC-iPSCs were used as a positive control, Scale bar, 50 mum. (D) Whole-genome sequencing of single-nucleotide variants (SNVs) in CS-fibroblasts, CS-iPSCs, GC-iPSCs, CS-MSCs and GC-MSCs. Sites with a heterozygosity percentage ranging between 0% and 30% were considered as SNV sites, and sites with a heterozygosity of >30% were considered as single-nucleotide polymorphisms (SNPs). (E) Whole-genome sequencing of copy number variations (CNVs) in CS-fibroblasts, CS-iPSCs, GC-iPSCs, CS-MSCs and GC-MSCs. Each point represents normalized coverage depth of each 500-kb genomic region of each chromosome. (F) Sterility and pathogen testing of the conditioned medium of GC-MSCs. a Endotoxin was identified as negative when the concentration was < 0.25 EU/mL. b CMV was identified as negative when the ratio of the OD 450 value of sample to the cut-off value (S/Co) was < 1.0. c HAV was identified as negative when the ratio of the cut-off value to the OD450 nm value of the s
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 2 Identification of human amnion mesenchymal stem cells. ( A ) Characteristics of passage three hAMSCs. ( B ) Osteogenic differentiation of hAMSCs indicated by Alizarin Red S. 100x. ( C ) Adipogenic differentiation of hAMSCs indicated by Oil Red O staining. 100x. ( D ) Surface antigens of hAMSCs were detected by flow cytometry. Cells were positive for CD90, CD105, and CD73 and negative for CD45, CD11b, CD34, and HLA-DR. ( E ) GFP-labeled hAMSCs and 293T cells were detected by fluorescence microscopy at day 3 following transfection with GFP-pseudovirion. 100x. ***p < 0.001.
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 1. Identification and characteristics of hPDLSCs. Morphological characterization of hPDLSCs in (A) primary culture and (B) culture at passage 3. A periodontal membrane fragment is present and shown in A. Flow cytometry analysis of surface markers expressed on hPDLSCs, showing that they were positive for (D) 0CD44, (F) CD90 and (G) CD105, and negative for (C) CD34 and (E) CD45. (H) Following 3 weeks of culture in osteogenic induction medium, the cells were stained with alizarin red. Mineralized nodules are shown. (I) Following 3 weeks of culture in adipogenic induction medium, the cells were stained with Oil Red O. Lipid globules are shown (black arrow). hPDLSCs, human periodontal ligament stem cells; PE, phycoerythrin.
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 2 ( A ) Results of fluorescence-activated cell sorting (FACS) analysis on hBM-MSCs surface markers (CD73 and CD105) and IgG control after PEMF for three days. ( B ) Immunohistochemical analysis of MAP-2 antibodies on hBM-MSCs cultured after PEMF for three days (original magnification: 100x); and immunofluorescence staining of NF-L antibodies on hBM-MSCs after PEMF for three days (original magnification: 400x).
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 1. Absence of ZKSCAN3 results in premature cellular senescence in hMSCs. ( A ) Western blot analysis of ZKSCAN3 protein in early-, middle- and late-passage (P3, P8, P13) hMSCs. Representative western blot images are shown on the left, and the statistical analysis of relative protein expression levels of ZKSCAN3 is on the right. GAPDH was used as the loading control. Data are presented as the mean +- SEMs, n = 3. * P < 0.05 (two tailed t -test). The asterisk indicates the band of ZKSCAN3 and P indicates passage. ( B ) Western blot analysis of ZKSCAN3 protein in LMNA +/+ , LMNA G608G/+ and LMNA G608G/G608G hMSCs (P7). Representative western blot images are shown on the left, and the statistical analysis of the relative protein expression levels of ZKSCAN3 is on the right. beta-actin was used as the loading control. Data are presented as the mean +- SEMs, n = 3. * P < 0.05, ** P < 0.01 (two tailed t -test). ( C ) Western blot analysis of ZKSCAN3 protein in WRN +/+ and WRN -/- hMSCs (P7). Representative western blot images are shown to the left, and the statistical analysis of the relative protein expression levels of ZKSCAN3 is on the right. beta-actin was used as the loading control. Data are presented as the mean +- SEMs, n = 3. ** P < 0.01 (two tailed t -test). ( D ) Western blot analysis of ZKSCAN3 protein in young and physiologically aged hMSCs (P7). Representative western blot images are shown on the left, and the statistical analysis of the relative protein express
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Characterization and uptake of SHED-Exos. A Surface markers of SHED cells were analyzed by flow cytometry (FACS) and were positive for mesenchymal markers (CD44, CD105 and CD90) and negative for endothelial markers (CD45, CD19 and CD14). B The morphology of exosomes (indicated by arrows) was observed using a transmission electron microscope (TEM). Scale bar = 100 nm. C Particle size distribution of SHED-Exos assessed by nanoparticle tracking analysis (NTA). D Expression of exosome-specific CD63 and TSG101 validated using western blotting. E Efficient uptake of PKH67-labeled exosomes by HUVECs was detected at 24 h. Scale bars = 100 mum
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 2. Endothelial progenitor cell characterization. Representative flow cytometry histograms of cEPCs for CD45, CD14, CD11b, CD105, CD115, and CD117 (A). n = 6 in each group of healthy control humans and patients exposed to SM.
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Decreased expression of CD90, CD73, and CD105 on Pre-T2D IPFP-ASCs. ( A ) Flow cytometry for CD90 and CD105 demonstrated decreased expression in Pre-T2D IPFP-ASCs compared to those from Non-T2D and T2D groups ( n = 3, * p < 0.05). ( B ) CFU assay illustrated increased self-renewal properties in T2D IPFP-ASCs compared to Non-T2D ( n = 3, * p < 0.05). ( C - F ) RT-qPCR for common adipokines in ASCs and adipocyte differentiated ASCs demonstrated no significant difference in adipogenic potential between Non-T2D, Pre-T2D, and T2D IPFP-ASCs ( n = 3, * p < 0.05, *** p < 0.001, **** p < 0.0001). Non-T2D IPFP ASCs 7-Day is the control Group set as 1. Non-T2D: IPFP-ASCs from donors without Type II diabetes mellitus, Pre-T2D: IPFP-ASCs from donors with pre-Type II diabetes mellitus, T2D: IPFP-ASCs from donors with type II diabetes mellitus, 7D: Confluent ASCs, AQ: Adipocyte Differentiated ASCs.
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure S2. Characterization of hiPSC-derived mesenchymal stem cells (MSCs). (A) The percentage of MSCs was determined at different time points by flow cytometry by detection of CD73 (ecto 5' nucleotidase), CD90 (Thy1), CD105 (Endoglin). (B) Representative flow cytometry analysis across samples to validate the absence of hematopoietic cell lineage markers CD34 and CD45. (C) After the first differentiation step, cells at 80% confluence were divided and expanded. The time between each passage is indicated. Cells were collected at P7, 40 d after differentiation for Hutchinson-Gilford progeria syndrome cells and 47 d for cells from the aged healthy donor. (D) The percentage of proliferating cells was monitored by flow cytometry at each passage using Ki67 staining. We observed a similar proliferation rate between the different conditions. Between each passage, measurements were made at three time points: 12, 24 and 48 h.
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 1 Primary human adipose-derived cells cultured in hypoxia (hAMSCs-H) and normoxia (hAMSCs-N) are both MSCs but normoxia-cultured cells show increased signs of senescence, such as increased area and elongated morphology, compared with hypoxia-cultured cells. ( a ) hAMSCs were isolated from human fat tissue and cultured in hypoxic (1.5% oxygen) or normoxic (21% oxygen) conditions in vitro . The viability, mobility, tumor tropism, safety, and tumorigenic potential were subsequently compared in vitro and in vivo . ( b ) Differentiation assay. hAMSCs were cultured in control media and differentiation media for 3 weeks, 10 days after the second passage. Three different stains were performed to assess differentiation capabilities (scale bar, 100 mu m). ( c ) Flow cytometric analysis was performed to confirm the absence of CD31-, CD34-, and CD45-positive cells in both cell cultures. In addition, primary hAMSC cultures expressed high levels of CD73, CD90, and CD105, both in hypoxic and normoxic culture conditions at day 10 after passage 2. ( d ) Representative images of cell morphologies of hAMSCs on 2D surface (scale bar, 200 mu m). ( e ) Schematic of 3D-nanopatterned surface used to assess morphology and motility. ( f ) Images of cell morphologies of hAMSCs on 3D-nanopatterned surface (scale bar, 200 mu m). ( g - j ) The length, width, area, and length-to-width ratio were measured and compared after cell aligned on the nanopattern surface. Error bars represent S.E.M. * P
- Conjugate
- Yellow dye
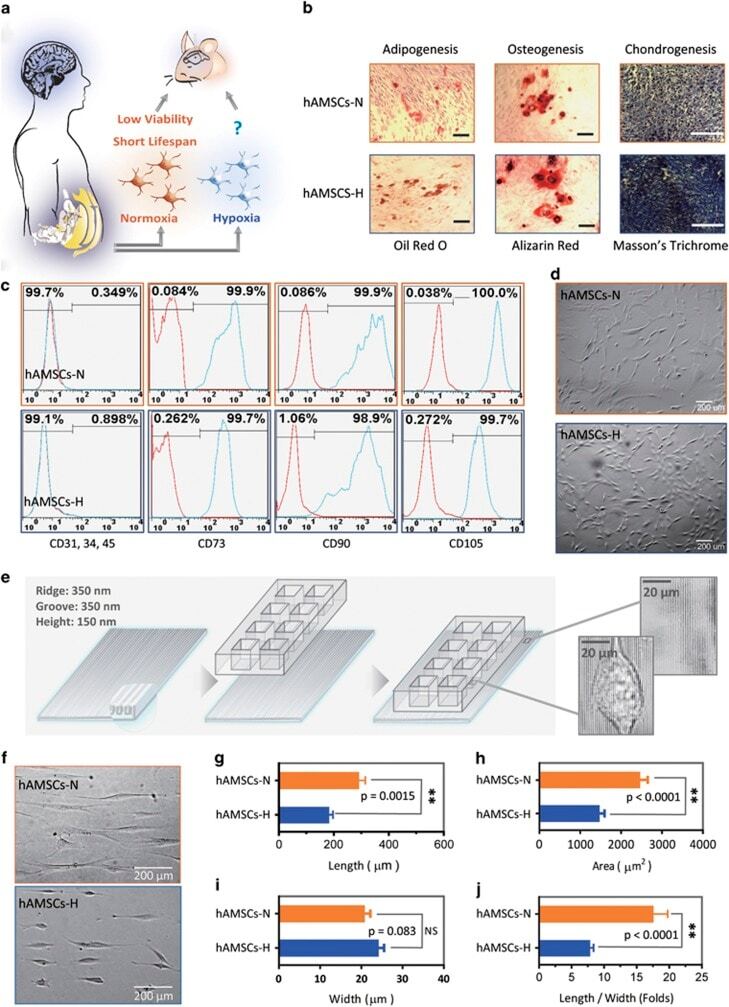
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 3 Hypoxia-cultured primary human adipose-derived mesenchymal stem cells (hAMSCs-H) retain a greater proliferation capacity compared with normoxia-cultured primary hAMSCs (hAMSCs-N) when exposed to GBM media. hAMSCs-H maintain stem cell characteristics when exposed to GBM media. ( a ) Representative MRI of GBM from a patient. ( b ) Schema showing the collection of GBM CM and culture of hAMSCs in filtered GBM CM for proliferation and migration assays. ( c ) MTT assay was used to determine the effects of hypoxic conditions on the proliferative capacity of primary hAMSCs in GBM CM. In GBM CM, hAMSCs-H showed greater proliferation at day 10 and 15 compared with hAMSCs-N. ( d ) Ki-67 immunostaining was performed to quantify the number of proliferating cells in GBM CM. Proliferative capacities of hAMSCs-H and hAMSCs-N are shown in GBM CM (normalized to hAMSC-N proliferative capacity in control media). In GBM CM, hAMSCs-H had a greater proportion of proliferating cells than hAMSCs-N. ( e) Differentiation assay. hAMSCs were cultured in control media, differentiation media, and GBM CM for 3 weeks, 10 days after the second passage. Three stainings were performed to assess the differentiation capabilities (scale bar, 100 mu m). Both hAMSCs-N and hAMSCs-H maintained tri-lineage differentiation capability in GBM CM. ( f ) Flow cytometric analysis for CD31, CD34, CD45, CD73, CD90, and CD105 in hAMSC-N and hAMSC-H cultures after exposure to GBM CM for 20 days. hAMSCs-H maintained MSC
- Conjugate
- Yellow dye
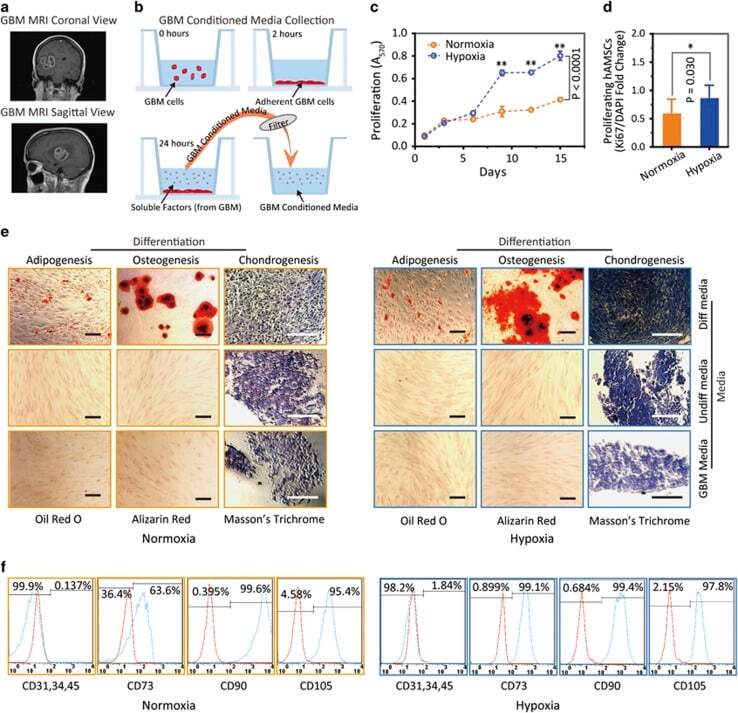
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 1 Characteristics of BM-MSCs. (a) Representative morphology of BM-MSCs. Scale bar = 500 mu m. (b) Representative flow cytometric characterization of cell surface markers expressed on BM-MSCs. Isotypic controls were represented by the gray filled histograms.
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 2 Characteristics of transfected BM-MSCs. (a) Representative morphology of transfected BM-MSCs in negative control group. (b) More than 90% of BM-MSCs expressed GFP in negative control group. (c) Representative morphology of transfected BM-MSCs in knock-down group. (d) More than 90% of BM-MSCs expressed GFP in knock-down group. (e) Representative flow cytometric characterization of cell surface markers expressed on transfected BM-MSCs. Isotypic controls were represented by black line. The red line represented the negative control group and the blue line represented the knock-down group. (f) The knock-down of NR2F2 was confirmed by western blot analysis.
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 2 CESCs Shared Features with BM-MSCs Regarding Morphology, Stem Cell Surface Markers, and Differentiation Ability (A) H&E staining of the tissue section. (B) Morphology of CESCs in agarose after seeding 6 weeks later. (C) Histologic section stained with Alcian blue of chondrified pellets in which CESCs formed in chondrogenic induction medium after 3 weeks. (D) Alizarin red staining of CESCs that underwent osteogenic induction for 3 weeks. (E) Immunophenotypic profile of stem cells in CESCs by flow cytometric analysis. The green lines indicate the fluorescence intensity of cells stained with the corresponding antibodies, and the red lines represent isotype-matched negative control cells. (F) Percentages of CESCs expressing different stem cell markers (n = 6 independent experiments). Data represent the mean +- SD.
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 3 Mesenchymal stem cell derivation. ( a ) MSC differentiation. Mono-layer iPSCs were subjected to bFGF, PDGFab and EGF resulting in differentiation to a cell population with spindle-shaped morphology (right). ( b ) FACS analysis. Passage 3 MSCs were analysed for cell surface expression of CD73, CD105, and CD90 ( n =3 experiments), and histogram analysis is shown in blue. Isotype antibody control FACS histograms are shown in pink. ( c , d ) Tri-lineage differentiation. ( c ) Oil red-O staining demonstrating the ability of iPSC-derived MSCs to form adipose cells. ( d ) Alizarin red staining of osteogenic progeny. ( e ) Toluidine blue staining of chondrogenic cells from MSCs. ( c - e ) Representative images of at least two different MSC pools and n =3-4 replicates. FACS, fluorescence-activated cell sorting.
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 1. Characterization of PDLSCs. (A) PDL cell clusters exhibited radiating or whirlpool-like morphology. The central structure in this image is a fragment of PDL tissue. Scale bar, 200 mum (B) CD146 + PDLSCs were small, round, fusiform and triangular. Scale bar, 100 mum. (C) PDLSCs were positive for the stem cell markers CD44, CD90 and CD105, but negative for CD34 and CD45, as detected by flow cytometry. PDL, periodontal ligament; PDLSCs, PDL stem cells.
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- 10.1371/journal.pbio.3000201.g002 Fig 2 YAP -/- hMSCs exhibit accelerated senescence. (A) Brightfield micrographs and FACS analysis of the surface markers CD105, CD73, and CD90 in WT, YAP -/- , and TAZ -/- hMSCs. Scale bar, 50 mum. (B) Cell growth curves of WT, YAP -/- , and TAZ -/- hMSCs. Data are presented as the mean +- SD, n = 3. (C) SA-beta-gal staining of WT, YAP -/- , and TAZ -/- hMSCs at passage 4. Scale bar, 100 mum. Data are presented as the mean +- SD, n = 3, *** P < 0.001. (D) Western blot analysis of YAP, TAZ, P16, P21, and P53 in WT, YAP -/- , and TAZ -/- hMSCs. GAPDH was used as a loading control (left). The protein levels normalized with GAPDH were shown as fold change relative to WT hMSCs (right). Data are presented as the mean +- SD, n = 3, * P < 0.05, ** P < 0.01. (E) Cell cycle analysis of WT, YAP -/- , and TAZ -/- hMSCs. Data are presented as the mean +- SD, n = 3, * P < 0.05, ** P < 0.01. (F) WT and YAP -/- hMSCs transduced with a lentivirus expressing Luc were injected into the TA muscle of immunodeficient mice. Luc activities were imaged at day (D)0, D1, D3, D5, and D7 after cell implantation. Representative images at D0 and D7 are shown. (G) Data are presented as the ratios of YAP -/- to WT cells (log 10 (fold)), mean +- SD, n = 5, * P < 0.05, *** P < 0.001. (H) SA-beta-gal staining of YAP -/- hMSCs transduced with lentiviruses expressing Luc or YAP. Data are presented as the mean +- SD, n = 3, *** P < 0. 001. (I) YAP -/- hMSCs overexpressing GFP plus
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 1 Growth characteristics of USCs. (a) The morphology of USCs by passage and differentiation. Single, small, compact rice grain-like cells were observed on the third day after initial seeding, and they formed a colony on the seventh day. The cells were considered to be at P0 when the confluence reached 70-80% and were passaged to the next generation. The USCs maintained the rice grain-like morphology after several passages, and USCs from the P3 generation were induced to differentiate into SMCs and UCs. The cells showed an elongated and spindle-shaped morphology after SMC differentiation and a cobblestone-shaped morphology after UC differentiation. Scale bar: 50 mu m, 100 mu m, and 200 mu m. (b) The growth curve of USCs from the P3 generation. (c) Detection of surface markers in USCs using flow cytometry. USCs did not express hematopoietic stem cell markers (CD31: 0.45%, CD34: 0.28%) but expressed MSC markers (CD44: 100%, CD73: 97.1%, and CD105: 96.3%) and pericyte markers (CD146: 95.7%). (d) Detection of surface markers in USCs using IF. USCs did not express hematopoietic stem cell markers (CD31, CD34, and CD45) but did express MSC markers (CD44 and CD133), the ESC marker SSEA4, and pericyte markers (CD146, PDGFRB, and NG2). NC: negative control; PDGFRB: platelet-derived growth factor beta-receptor; NG2: neural/glial antigen 2. Scale bar: 25 mu m.
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Fig. 2 DGCR8 dex2 accelerates cellular senescence in hMSCs. a FACS analysis of the MSC markers CD73, CD90, and CD105 in WT and DR8 dex2 hMSCs. b Western blotting analysis of DGCR8 in WT and DR8 dex2 hMSCs. beta-actin was used as the loading control. c Growth curve of WT and DR8 dex2 hMSCs. d Clonal expansion analysis of WT and DR8 dex2 hMSCs at early passage (P3) and late passage (P6). Scale bar, 30 mum. Data were presented as mean +- SEM. n = 3 wells per cell type, *** p < 0.001. e SA-beta-gal staining of WT and DR8 dex2 hMSCs at early (P3) and late (P6) passages. Scale bar, 20 mum. Data were presented as mean +- SEM. n = 4 images per cell type, *** p < 0.001. f Western blotting analysis of P16, P21, and GATA4 in WT and DR8 dex2 hMSCs. beta-actin was used as the loading control. g Heatmap showing quantitative RT-PCR analysis of the indicated genes in P3- and P6-hMSCs. Expression levels of the indicated genes in DR8 dex2 hMSCs were normalized to those in WT hMSCs. ( h ) Whole genome analysis of copy number variations (CNVs) in WT and DR8 dex2 hMSCs. i Photon flux from tibialis anterior (TA) muscles of nude mice transplanted with WT (left) or DR8 dex2 hMSCs (right) that express luciferase. The attrition of hMSCs after implantation was measured by the reduction of luciferase activity in TA muscles using an in vivo imaging system (IVIS). Data were presented as mean +- SEM. n = 3 mice per group, * p < 0.05. j SA-beta-gal staining of DR8 dex2
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 1. Characterization of hUC-MSCs. (A) Morphological observations of hUC-MSCs. Umbilical cord tissues were cultured for >15 days and long spindle-shaded fibroblastic cells were observed around the tissue using Zeiss light microscopy (scale bar, 100 um). (B) Phenotyping of hUC-MSCs. hUC-MSCs were stained with a fluorescein-labeled antibodies (CD34, CD45, CD73, CD90, CD105, CD14, CD19 and HLA-DR) and analyzed with a flow cytometer. (C) Adipogenic and (D) osteogenic differentiation of hUC-MSCs. hUC-MSCs were cultured in adipogenic and osteogenic medium, respectively. Lipid droplets in the adipocytes are presented with Oil Red O staining (scale bar, 100 um). hUC-MSCs-derived osteoblasts were detected with Alizarin Red staining (scale bar, 200 um). (E) hUC-MSCs inhibit the proliferation of CFSE-labeled CD4 + T cells, which were activated by Con A stimulation. Experiments were repeated three times and representative graphs and images are presented. hUC-MSC, human umbilical cord-derived mesenchymal stem cell; MSC Sup, culture supernatant of hUC-MSCs; Con A, concanavalin A; CFSE, carboxyfluorescein succinimidyl ester.
- Conjugate
- Yellow dye
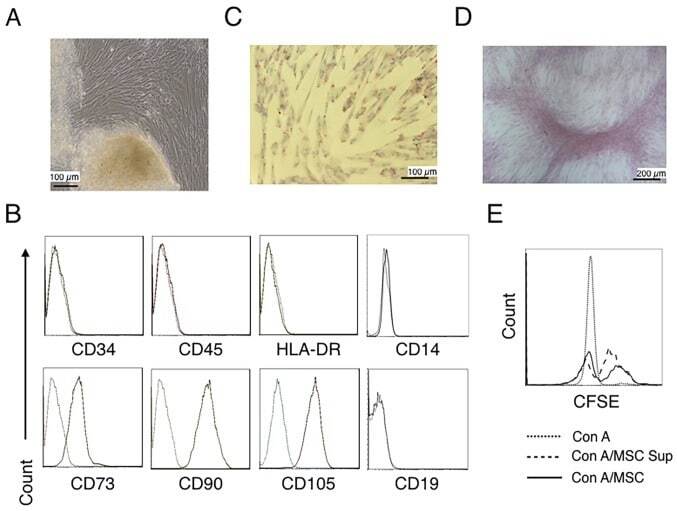
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 2 RAP1 -/- hMSCs exhibited retarded cellular senescence. (A) Brightfield micrographs of hMSCs showed normal morphology. Scale bar, 50 um. (B) Flow cytometry demonstrated that sorted hMSCs uniformly expressed the MSC-specific surface markers CD73, CD90 and CD105. (C) qRT-PCR analysis demonstrated the deletion of RAP1 at the transcriptional level in RAP1 -/- hMSCs by primers P8 and P9. Data were presented as the mean +- SEM, n = 3. *** P < 0.001. (D) Immunofluorescence micrographs of RAP1 in WT and RAP1 -/- hMSCs. Scale bar, 10 um. (E) Western blotting analysis demonstrated the absence of RAP1 in RAP1 -/- hMSCs. beta-Actin was used as a loading control. (F) Characterization of the differentiation potential of hMSCs. Toluidine blue O, Oil Red O and Von Kossa staining were used to detect chondrocytes, adipocytes and osteoblasts, respectively. Scale bar, 200 um. (G) Cell growth curves showed the enhanced proliferation ability of RAP1 -/- hMSCs. (H) Immunostaining of the proliferation marker Ki67 in WT and RAP1 -/- hMSCs. EP (early passage) and LP (late passage) represented P2 and P9, respectively. Scale bar, 50 um. Data were presented as the mean +- SEM, n = 6. NS, not significant, *** P < 0.001. (I) Clonal expansion analysis of WT and RAP1 -/- hMSCs. EP and LP represented P2 and P9, respectively. Scale bar, 50 um. Data were presented as the mean +- SEM, n = 3. ** P < 0.01, *** P < 0.001. (J) Cell cycle analysis of WT and RAP1 -/- hMSCs (LP). Data
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Fig. 3. The phenotype of MenSCs. (A,B) To determine the immunophenotype of MenSCs, P3 MenSCs-DGC ( n =5) and MenSCs-RLB ( n =5) were stained by corresponding conjugated antibodies and analyzed by FACS. Both MenSCs-DGC and MenSCs-RLB positively expressed classical ASCs' markers (CD29, CD44, CD73, CD90 and CD105) and HLA-ABC; they did not express hematopoietic stem cell markers (CD34 and CD45) and HLA-DR. (C) The quantification of flow cytometry results of A and B.
- Conjugate
- Yellow dye
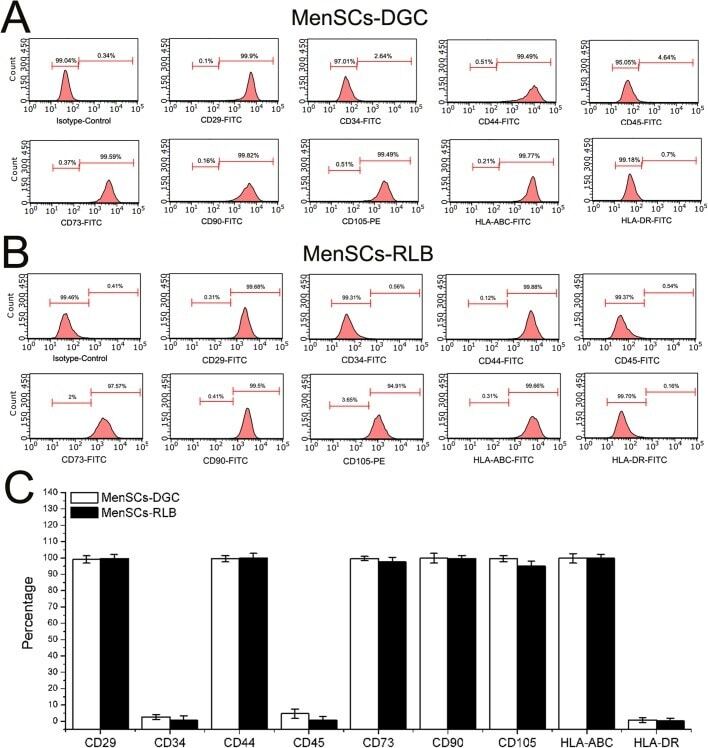
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 2 Flow cytometry of CD cell surface markers for cells cultured under hypoxia and normoxia. The positive CD markers for MSCs as detected by the fluorescent antibodies anti-CD73 FITC, anti-CD105 PE, and anti-CD90 PE Cy7. The negative markers of MSCs were detected using anti-CD14 FITC, anti-CD45 PerCP, anti-CD34-R-PE, and anti-CD19 PE-Cy7 antibodies. Unstained cell for each condition was used as negative controls.
- Conjugate
- Yellow dye

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 1 Human amniotic fluid-derived mesenchymal stem cells (AF-MSCs) characterization. ( A ) The typical morphology of human amniotic fluid-derived mesenchymal stem cells, grown in cell culture. Scale bar = 400 um. ( B ) The expression of the main cell surface markers CD44, CD90, CD105, and CD34 as detected by flow cytometry. Unlabeled ctrl: unlabeled, undifferentiated control cells. Results are presented as the mean +- SD ( n = 3). ( C ) The relative expression of pluripotency gene markers, namely, OCT4, SOX2, NANOG, and REX1, as determined by RT-qPCR. Data, relative to GAPDH, are presented as the mean +- SD ( n = 3).
- Conjugate
- Yellow dye
