Explore
Explore Validate
Validate Learn
Learn Western blot
Western blot Immunoprecipitation
ImmunoprecipitationAntibody data
- Antibody Data
- Antigen structure
- References [19]
- Comments [0]
- Validations
- Western blot [2]
- Immunohistochemistry [2]
Submit
Validation data
Reference
Comment
Report error
- Product number
- AF933 - Provider product page

- Provider
- R&D Systems
- Product name
- Human/Mouse/Rat/Hamster ACE-2 Antibody
- Antibody type
- Polyclonal
- Description
- Antigen Affinity-purified. Detects human ACE-2 in direct ELISAs. Detects human, mouse, and rat ACE-2 in Western blots. Detects Hamster ACE-2 in immunohistochemistry. In direct ELISAs and Western blots, less than 1% cross-reactivity with recombinant human ACE is observed.
- Reactivity
- Human, Mouse, Rat, Hamster
- Host
- Goat
- Conjugate
- Unconjugated
- Antigen sequence
Q9BYF1- Isotype
- IgG
- Vial size
- 100 ug
- Concentration
- LYOPH
- Storage
- Use a manual defrost freezer and avoid repeated freeze-thaw cycles. 12 months from date of receipt, -20 to -70 °C as supplied. 1 month, 2 to 8 °C under sterile conditions after reconstitution. 6 months, -20 to -70 °C under sterile conditions after reconstitution.
Submitted references Translation of Angiotensin-Converting Enzyme 2 upon Liver- and Lung-Targeted Delivery of Optimized Chemically Modified mRNA.
Protein Kinase C-δ Mediates Shedding of Angiotensin-Converting Enzyme 2 from Proximal Tubular Cells.
HTCC: Broad Range Inhibitor of Coronavirus Entry.
Surface vimentin is critical for the cell entry of SARS-CoV.
Characterization of angiotensin-converting enzyme 2 ectodomain shedding from mouse proximal tubular cells.
TMPRSS2 and ADAM17 cleave ACE2 differentially and only proteolysis by TMPRSS2 augments entry driven by the severe acute respiratory syndrome coronavirus spike protein.
Isolation and characterization of a bat SARS-like coronavirus that uses the ACE2 receptor.
In-vitro renal epithelial cell infection reveals a viral kidney tropism as a potential mechanism for acute renal failure during Middle East Respiratory Syndrome (MERS) Coronavirus infection.
Influenza and SARS-coronavirus activating proteases TMPRSS2 and HAT are expressed at multiple sites in human respiratory and gastrointestinal tracts.
Intrarenal renin angiotensin system revisited: role of megalin-dependent endocytosis along the proximal nephron.
Importance of cholesterol-rich membrane microdomains in the interaction of the S protein of SARS-coronavirus with the cellular receptor angiotensin-converting enzyme 2.
Lipid rafts are involved in SARS-CoV entry into Vero E6 cells.
Difference in receptor usage between severe acute respiratory syndrome (SARS) coronavirus and SARS-like coronavirus of bat origin.
Detection of soluble angiotensin-converting enzyme 2 in heart failure: insights into the endogenous counter-regulatory pathway of the renin-angiotensin-aldosterone system.
Synthetic reconstruction of zoonotic and early human severe acute respiratory syndrome coronavirus isolates that produce fatal disease in aged mice.
Clathrin-dependent entry of severe acute respiratory syndrome coronavirus into target cells expressing ACE2 with the cytoplasmic tail deleted.
Interferon-gamma and interleukin-4 downregulate expression of the SARS coronavirus receptor ACE2 in Vero E6 cells.
Apical entry and release of severe acute respiratory syndrome-associated coronavirus in polarized Calu-3 lung epithelial cells.
Susceptibility to SARS coronavirus S protein-driven infection correlates with expression of angiotensin converting enzyme 2 and infection can be blocked by soluble receptor.
Schrom E, Huber M, Aneja M, Dohmen C, Emrich D, Geiger J, Hasenpusch G, Herrmann-Janson A, Kretzschmann V, Mykhailyk O, Pasewald T, Oak P, Hilgendorff A, Wohlleber D, Hoymann HG, Schaudien D, Plank C, Rudolph C, Kubisch-Dohmen R
Molecular therapy. Nucleic acids 2017 Jun 16;7:350-365
Molecular therapy. Nucleic acids 2017 Jun 16;7:350-365
Protein Kinase C-δ Mediates Shedding of Angiotensin-Converting Enzyme 2 from Proximal Tubular Cells.
Xiao F, Zimpelmann J, Burger D, Kennedy C, Hébert RL, Burns KD
Frontiers in pharmacology 2016;7:146
Frontiers in pharmacology 2016;7:146
HTCC: Broad Range Inhibitor of Coronavirus Entry.
Milewska A, Kaminski K, Ciejka J, Kosowicz K, Zeglen S, Wojarski J, Nowakowska M, Szczubiałka K, Pyrc K
PloS one 2016;11(6):e0156552
PloS one 2016;11(6):e0156552
Surface vimentin is critical for the cell entry of SARS-CoV.
Yu YT, Chien SC, Chen IY, Lai CT, Tsay YG, Chang SC, Chang MF
Journal of biomedical science 2016 Jan 22;23:14
Journal of biomedical science 2016 Jan 22;23:14
Characterization of angiotensin-converting enzyme 2 ectodomain shedding from mouse proximal tubular cells.
Xiao F, Zimpelmann J, Agaybi S, Gurley SB, Puente L, Burns KD
PloS one 2014;9(1):e85958
PloS one 2014;9(1):e85958
TMPRSS2 and ADAM17 cleave ACE2 differentially and only proteolysis by TMPRSS2 augments entry driven by the severe acute respiratory syndrome coronavirus spike protein.
Heurich A, Hofmann-Winkler H, Gierer S, Liepold T, Jahn O, Pöhlmann S
Journal of virology 2014 Jan;88(2):1293-307
Journal of virology 2014 Jan;88(2):1293-307
Isolation and characterization of a bat SARS-like coronavirus that uses the ACE2 receptor.
Ge XY, Li JL, Yang XL, Chmura AA, Zhu G, Epstein JH, Mazet JK, Hu B, Zhang W, Peng C, Zhang YJ, Luo CM, Tan B, Wang N, Zhu Y, Crameri G, Zhang SY, Wang LF, Daszak P, Shi ZL
Nature 2013 Nov 28;503(7477):535-8
Nature 2013 Nov 28;503(7477):535-8
In-vitro renal epithelial cell infection reveals a viral kidney tropism as a potential mechanism for acute renal failure during Middle East Respiratory Syndrome (MERS) Coronavirus infection.
Eckerle I, Müller MA, Kallies S, Gotthardt DN, Drosten C
Virology journal 2013 Dec 23;10:359
Virology journal 2013 Dec 23;10:359
Influenza and SARS-coronavirus activating proteases TMPRSS2 and HAT are expressed at multiple sites in human respiratory and gastrointestinal tracts.
Bertram S, Heurich A, Lavender H, Gierer S, Danisch S, Perin P, Lucas JM, Nelson PS, Pöhlmann S, Soilleux EJ
PloS one 2012;7(4):e35876
PloS one 2012;7(4):e35876
Intrarenal renin angiotensin system revisited: role of megalin-dependent endocytosis along the proximal nephron.
Pohl M, Kaminski H, Castrop H, Bader M, Himmerkus N, Bleich M, Bachmann S, Theilig F
The Journal of biological chemistry 2010 Dec 31;285(53):41935-46
The Journal of biological chemistry 2010 Dec 31;285(53):41935-46
Importance of cholesterol-rich membrane microdomains in the interaction of the S protein of SARS-coronavirus with the cellular receptor angiotensin-converting enzyme 2.
Glende J, Schwegmann-Wessels C, Al-Falah M, Pfefferle S, Qu X, Deng H, Drosten C, Naim HY, Herrler G
Virology 2008 Nov 25;381(2):215-21
Virology 2008 Nov 25;381(2):215-21
Lipid rafts are involved in SARS-CoV entry into Vero E6 cells.
Lu Y, Liu DX, Tam JP
Biochemical and biophysical research communications 2008 May 2;369(2):344-9
Biochemical and biophysical research communications 2008 May 2;369(2):344-9
Difference in receptor usage between severe acute respiratory syndrome (SARS) coronavirus and SARS-like coronavirus of bat origin.
Ren W, Qu X, Li W, Han Z, Yu M, Zhou P, Zhang SY, Wang LF, Deng H, Shi Z
Journal of virology 2008 Feb;82(4):1899-907
Journal of virology 2008 Feb;82(4):1899-907
Detection of soluble angiotensin-converting enzyme 2 in heart failure: insights into the endogenous counter-regulatory pathway of the renin-angiotensin-aldosterone system.
Epelman S, Tang WH, Chen SY, Van Lente F, Francis GS, Sen S
Journal of the American College of Cardiology 2008 Aug 26;52(9):750-4
Journal of the American College of Cardiology 2008 Aug 26;52(9):750-4
Synthetic reconstruction of zoonotic and early human severe acute respiratory syndrome coronavirus isolates that produce fatal disease in aged mice.
Rockx B, Sheahan T, Donaldson E, Harkema J, Sims A, Heise M, Pickles R, Cameron M, Kelvin D, Baric R
Journal of virology 2007 Jul;81(14):7410-23
Journal of virology 2007 Jul;81(14):7410-23
Clathrin-dependent entry of severe acute respiratory syndrome coronavirus into target cells expressing ACE2 with the cytoplasmic tail deleted.
Inoue Y, Tanaka N, Tanaka Y, Inoue S, Morita K, Zhuang M, Hattori T, Sugamura K
Journal of virology 2007 Aug;81(16):8722-9
Journal of virology 2007 Aug;81(16):8722-9
Interferon-gamma and interleukin-4 downregulate expression of the SARS coronavirus receptor ACE2 in Vero E6 cells.
de Lang A, Osterhaus AD, Haagmans BL
Virology 2006 Sep 30;353(2):474-81
Virology 2006 Sep 30;353(2):474-81
Apical entry and release of severe acute respiratory syndrome-associated coronavirus in polarized Calu-3 lung epithelial cells.
Tseng CT, Tseng J, Perrone L, Worthy M, Popov V, Peters CJ
Journal of virology 2005 Aug;79(15):9470-9
Journal of virology 2005 Aug;79(15):9470-9
Susceptibility to SARS coronavirus S protein-driven infection correlates with expression of angiotensin converting enzyme 2 and infection can be blocked by soluble receptor.
Hofmann H, Geier M, Marzi A, Krumbiegel M, Peipp M, Fey GH, Gramberg T, Pöhlmann S
Biochemical and biophysical research communications 2004 Jul 9;319(4):1216-21
Biochemical and biophysical research communications 2004 Jul 9;319(4):1216-21
No comments: Submit comment
Supportive validation
- Submitted by
- R&D Systems (provider)
- Main image
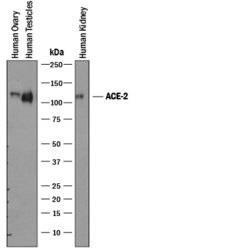
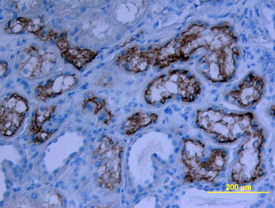
- Experimental details
- Detection of Human ACE-2 by Western Blot. Western blot shows lysates of human ovary tissue, human testis tissue, and human kidney tissue. PVDF membrane was probed with 1 µg/mL of Goat Anti-Human ACE-2 Antigen Affinity-purified Polyclonal Antibody (Catalog # AF933) followed by HRP-conjugated Anti-Goat IgG Secondary Antibody (Catalog # HAF019). A specific band was detected for ACE-2 at approximately 120 kDa (as indicated). This experiment was conducted under reducing conditions and using Immunoblot Buffer Group 1.

- Submitted by
- R&D Systems (provider)
- Main image

- Experimental details
- Detection of Human ACE-2 by Simple WesternTM. Simple Western lane view shows lysates of human kidney tissue, loaded at 0.2 mg/mL. A specific band was detected for ACE-2 at approximately 155 kDa (as indicated) using 10 µg/mL of Goat Anti-Human ACE-2 Antigen Affinity-purified Polyclonal Antibody (Catalog # AF933) followed by 1:50 dilution of HRP-conjugated Anti-Goat IgG Secondary Antibody (Catalog # HAF109). This experiment was conducted under reducing conditions and using the 12-230 kDa separation system.

Supportive validation
- Submitted by
- R&D Systems (provider)
- Main image
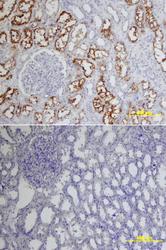
- Experimental details
- ACE-2 in Human Kidney. ACE-2 was detected in immersion fixed paraffin-embedded sections of human kidney using Goat Anti-Human ACE-2 Antigen Affinity-purified Polyclonal Antibody (Catalog # AF933) at 15 µg/mL overnight at 4 °C. Tissue was stained using the Anti-Goat HRP-DAB Cell & Tissue Staining Kit (brown; Catalog # CTS008) and counterstained with hematoxylin (blue). Lower panel shows a lack of labeling if primary antibodies are omitted and tissue is stained only with secondary antibody followed by incubation with detection reagents. View our protocol for Chromogenic IHC Staining of Paraffin-embedded Tissue Sections.

- Submitted by
- R&D Systems (provider)
- Main image
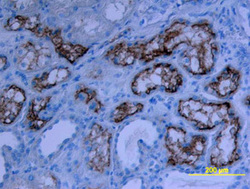
- Experimental details
- ACE-2 in Human Kidney. ACE-2 was detected in immersion fixed paraffin-embedded sections of human kidney using Goat Anti-Human ACE-2 Antigen Affinity-purified Polyclonal Antibody (Catalog # AF933) at 15 µg/mL overnight at 4 °C. Tissue was stained using the Anti-Goat HRP-DAB Cell & Tissue Staining Kit (brown; Catalog # CTS008) and counterstained with hematoxylin (blue). View our protocol for Chromogenic IHC Staining of Paraffin-embedded Tissue Sections.