Explore
Explore Validate
Validate Learn
LearnMA3-920
antibody from Invitrogen Antibodies
Targeting: CACNA1S
CACNL1A3, Cav1.1, HOKPP, hypoPP, MHS5
 Western blot
Western blot Immunohistochemistry
ImmunohistochemistryAntibody data
- Antibody Data
- Antigen structure
- References [79]
- Comments [0]
- Validations
- Immunohistochemistry [1]
- Flow cytometry [1]
- Other assay [26]
Submit
Validation data
Reference
Comment
Report error
- Product number
- MA3-920 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- CaV1.1 Monoclonal Antibody (1A)
- Antibody type
- Monoclonal
- Antigen
- Purifed from natural sources
- Description
- MA3-920 detects 1,4-dihydropyridine (DHP) receptor alpha-1 subunit in human, rat, mouse, guinea pig and rabbit skeletal muscle. The DHP Receptor alpha-1 protein is also known as CACNA1S or Cav1.1 alpha-1 subunit. MA3-920 has been successfully used in Western blot, FACS, immunohistochemistry and immunoprecipitation procedures. By Western blot, this antibody detects an ~200 kDa protein representing the DHP receptor in rat skeletal muscle extracts. Immunohistochemical staining of DHP receptor in rabbit skeletal muscle with MA3-920 results in double rows of discrete punctate staining representing pairs of triads on the opposing sides of the Z-lines. This product can be used to inhibit the DHP-sensitive calcium current in BC3H1 mouse muscle cells. The MA3-920 antigen is purified rabbit muscle T-tubule DHP receptor.
- Reactivity
- Human, Mouse, Rat, Guinea Pig, Rabbit
- Host
- Mouse
- Isotype
- IgG
- Antibody clone number
- 1A
- Vial size
- 200 µL
- Concentration
- 1 mg/mL
- Storage
- -20° C, Avoid Freeze/Thaw Cycles
Submitted references Myogenesis modelled by human pluripotent stem cells: a multi-omic study of Duchenne myopathy early onset.
Differential physiological roles for BIN1 isoforms in skeletal muscle development, function and regeneration.
Functional skeletal muscle model derived from SOD1-mutant ALS patient iPSCs recapitulates hallmarks of disease progression.
Ca(2+)-activated Cl(-) channel TMEM16A/ANO1 identified in zebrafish skeletal muscle is crucial for action potential acceleration.
Maximal strength training increases muscle force generating capacity and the anaerobic ATP synthesis flux without altering the cost of contraction in elderly.
Voltage-dependent Ca(2+) channels promote branching morphogenesis of salivary glands by patterning differential growth.
l-arginine ingestion inhibits eccentric contraction-induced proteolysis and force deficit via S-nitrosylation of calpain.
Sarcolipin deletion exacerbates soleus muscle atrophy and weakness in phospholamban overexpressing mice.
A chemical chaperone improves muscle function in mice with a RyR1 mutation.
Calcium dysregulation, functional calpainopathy, and endoplasmic reticulum stress in sporadic inclusion body myositis.
Dysferlin mediates membrane tubulation and links T-tubule biogenesis to muscular dystrophy.
Congenital myopathy results from misregulation of a muscle Ca2+ channel by mutant Stac3.
Role of calpain in eccentric contraction-induced proteolysis of Ca(2+)-regulatory proteins and force depression in rat fast-twitch skeletal muscle.
Amphiphysin (BIN1) negatively regulates dynamin 2 for normal muscle maturation.
The SH3 and cysteine-rich domain 3 (Stac3) gene is important to growth, fiber composition, and calcium release from the sarcoplasmic reticulum in postnatal skeletal muscle.
Thrombospondin expression in myofibers stabilizes muscle membranes.
Alternative Splicing of Four Trafficking Genes Regulates Myofiber Structure and Skeletal Muscle Physiology.
Genetic overexpression of Serpina3n attenuates muscular dystrophy in mice.
Eps 15 Homology Domain (EHD)-1 Remodels Transverse Tubules in Skeletal Muscle.
Inward rectifier potassium currents in mammalian skeletal muscle fibres.
Downregulation of connexin43 by microRNA-130a in cardiomyocytes results in cardiac arrhythmias.
EHD1 mediates vesicle trafficking required for normal muscle growth and transverse tubule development.
Dysferlin and myoferlin regulate transverse tubule formation and glycerol sensitivity.
Gene therapy prolongs survival and restores function in murine and canine models of myotubular myopathy.
Perturbations in intracellular Ca2+ handling in skeletal muscle in the G93A*SOD1 mouse model of amyotrophic lateral sclerosis.
Effects of SEA0400 on arrhythmogenicity in a Langendorff-perfused 1-month myocardial infarction rabbit model.
Deregulation of the protocadherin gene FAT1 alters muscle shapes: implications for the pathogenesis of facioscapulohumeral dystrophy.
Dysferlin stabilizes stress-induced Ca2+ signaling in the transverse tubule membrane.
Lack of myotubularin (MTM1) leads to muscle hypotrophy through unbalanced regulation of the autophagy and ubiquitin-proteasome pathways.
Highly conductive carbon nanotube matrix accelerates developmental chloride extrusion in central nervous system neurons by increased expression of chloride transporter KCC2.
Domain cooperativity in the β1a subunit is essential for dihydropyridine receptor voltage sensing in skeletal muscle.
Myotubularin and PtdIns3P remodel the sarcoplasmic reticulum in muscle in vivo.
Abnormalities of calcium handling proteins in skeletal muscle mirror those of the heart in humans with heart failure: a shared mechanism?
Muscle weakness in myotonic dystrophy associated with misregulated splicing and altered gating of Ca(V)1.1 calcium channel.
Three-dimensional localization of the α and β subunits and of the II-III loop in the skeletal muscle L-type Ca2+ channel.
Increased expression of wild-type or a centronuclear myopathy mutant of dynamin 2 in skeletal muscle of adult mice leads to structural defects and muscle weakness.
Identification of novel ryanodine receptor 1 (RyR1) protein interaction with calcium homeostasis endoplasmic reticulum protein (CHERP).
The calcium channel α2/δ1 subunit interacts with ATP5b in the plasma membrane of developing muscle cells.
Aquaporin-4 water channel oligomers are associated with the transverse tubules of skeletal myofibers.
Defects in amphiphysin 2 (BIN1) and triads in several forms of centronuclear myopathies.
Orthograde dihydropyridine receptor signal regulates ryanodine receptor passive leak.
Aerobic exercise training improves skeletal muscle function and Ca2+ handling-related protein expression in sympathetic hyperactivity-induced heart failure.
Initiating treadmill training in late middle age offers modest adaptations in Ca2+ handling but enhances oxidative damage in senescent rat skeletal muscle.
Calsequestrin and junctin immunoreactivity in hexagonally cross-linked tubular arrays myopathy.
Aerobic exercise training improves Ca2+ handling and redox status of skeletal muscle in mice.
Skeletal muscle type comparison of subsarcolemmal mitochondrial membrane phospholipid fatty acid composition in rat.
T-tubule disorganization and defective excitation-contraction coupling in muscle fibers lacking myotubularin lipid phosphatase.
Exercise-induced regulation of phospholemman (FXYD1) in rat skeletal muscle: implications for Na+/K+-ATPase activity.
Reduced expression of sarcalumenin and related Ca2+ -regulatory proteins in aged rat skeletal muscle.
Zebrafish relatively relaxed mutants have a ryanodine receptor defect, show slow swimming and provide a model of multi-minicore disease.
Excitation-contraction coupling in airway smooth muscle.
A Ca2+-binding domain in RyR1 that interacts with the calmodulin binding site and modulates channel activity.
Neuregulin receptor ErbB2 localization at T-tubule in cardiac and skeletal muscle.
Restricted distribution of mRNAs encoding a sarcoplasmic reticulum or transverse tubule protein in skeletal myofibers.
Developmental induction of DHPR alpha 1s and RYR1 gene expression does not require neural or mechanical signals.
Regulation of dihydropyridine receptor gene expression in mouse skeletal muscles by stretch and disuse.
Increased sensitivity of the ryanodine receptor to halothane-induced oligomerization in malignant hyperthermia-susceptible human skeletal muscle.
Na,K-ATPase alpha- and beta-isoform expression in developing skeletal muscles: alpha(2) correlates with t-tubule formation.
Homer proteins and InsP(3) receptors co-localise in the longitudinal sarcoplasmic reticulum of skeletal muscle fibres.
Homer proteins and InsP(3) receptors co-localise in the longitudinal sarcoplasmic reticulum of skeletal muscle fibres.
Sarcolemmal organization in skeletal muscle lacking desmin: evidence for cytokeratins associated with the membrane skeleton at costameres.
Suramin interacts with the calmodulin binding site on the ryanodine receptor, RYR1.
Agonists cause endocytosis of nicotinic acetylcholine receptors on cultured myotubes.
Ca2+-induced Ca2+ release supports the relay mode of activity in thalamocortical cells.
Ca2+-induced Ca2+ release supports the relay mode of activity in thalamocortical cells.
Extensive but coordinated reorganization of the membrane skeleton in myofibers of dystrophic (mdx) mice.
Cloning of a calcium channel alpha1 subunit from the reef-building coral, Stylophora pistillata.
A transgenic myogenic cell line lacking ryanodine receptor protein for homologous expression studies: reconstitution of Ry1R protein and function.
A transgenic myogenic cell line lacking ryanodine receptor protein for homologous expression studies: reconstitution of Ry1R protein and function.
Dyspedic mouse skeletal muscle expresses major elements of the triadic junction but lacks detectable ryanodine receptor protein and function.
Dyspedic mouse skeletal muscle expresses major elements of the triadic junction but lacks detectable ryanodine receptor protein and function.
Branching points of renal resistance arteries are enriched in L-type calcium channels and initiate vasoconstriction.
Branching points of renal resistance arteries are enriched in L-type calcium channels and initiate vasoconstriction.
Alpha 1 and alpha 2 Ca2+ channel subunit expression in human neuronal and small cell carcinoma cells.
Triad formation: organization and function of the sarcoplasmic reticulum calcium release channel and triadin in normal and dysgenic muscle in vitro.
Localization of the alpha 1 and alpha 2 subunits of the dihydropyridine receptor and ankyrin in skeletal muscle triads.
The alpha 1 and alpha 2 polypeptides of the dihydropyridine-sensitive calcium channel differ in developmental expression and tissue distribution.
Monoclonal antibody to the alpha 1-subunit of the dihydropyridine-binding complex inhibits calcium currents in BC3H1 myocytes.
Monoclonal antibody identifies a 200-kDa subunit of the dihydropyridine-sensitive calcium channel.
Mournetas V, Massouridès E, Dupont JB, Kornobis E, Polvèche H, Jarrige M, Dorval ARL, Gosselin MRF, Manousopoulou A, Garbis SD, Górecki DC, Pinset C
Journal of cachexia, sarcopenia and muscle 2021 Feb;12(1):209-232
Journal of cachexia, sarcopenia and muscle 2021 Feb;12(1):209-232
Differential physiological roles for BIN1 isoforms in skeletal muscle development, function and regeneration.
Prokic I, Cowling BS, Kutchukian C, Kretz C, Tasfaout H, Gache V, Hergueux J, Wendling O, Ferry A, Toussaint A, Gavriilidis C, Nattarayan V, Koch C, Lainé J, Combe R, Tiret L, Jacquemond V, Pilot-Storck F, Laporte J
Disease models & mechanisms 2020 Nov 24;13(11)
Disease models & mechanisms 2020 Nov 24;13(11)
Functional skeletal muscle model derived from SOD1-mutant ALS patient iPSCs recapitulates hallmarks of disease progression.
Badu-Mensah A, Guo X, McAleer CW, Rumsey JW, Hickman JJ
Scientific reports 2020 Aug 31;10(1):14302
Scientific reports 2020 Aug 31;10(1):14302
Ca(2+)-activated Cl(-) channel TMEM16A/ANO1 identified in zebrafish skeletal muscle is crucial for action potential acceleration.
Dayal A, Ng SFJ, Grabner M
Nature communications 2019 Jan 10;10(1):115
Nature communications 2019 Jan 10;10(1):115
Maximal strength training increases muscle force generating capacity and the anaerobic ATP synthesis flux without altering the cost of contraction in elderly.
Berg OK, Kwon OS, Hureau TJ, Clifton HL, Thurston T, Le Fur Y, Jeong EK, Amann M, Richardson RS, Trinity JD, Wang E, Layec G
Experimental gerontology 2018 Oct 1;111:154-161
Experimental gerontology 2018 Oct 1;111:154-161
Voltage-dependent Ca(2+) channels promote branching morphogenesis of salivary glands by patterning differential growth.
Kim JM, Choi S, Lee SW, Park K
Scientific reports 2018 May 15;8(1):7566
Scientific reports 2018 May 15;8(1):7566
l-arginine ingestion inhibits eccentric contraction-induced proteolysis and force deficit via S-nitrosylation of calpain.
Kanzaki K, Watanabe D, Aibara C, Kawakami Y, Yamada T, Takahashi Y, Wada M
Physiological reports 2018 Jan;6(2)
Physiological reports 2018 Jan;6(2)
Sarcolipin deletion exacerbates soleus muscle atrophy and weakness in phospholamban overexpressing mice.
Fajardo VA, Gamu D, Mitchell A, Bloemberg D, Bombardier E, Chambers PJ, Bellissimo C, Quadrilatero J, Tupling AR
PloS one 2017;12(3):e0173708
PloS one 2017;12(3):e0173708
A chemical chaperone improves muscle function in mice with a RyR1 mutation.
Lee CS, Hanna AD, Wang H, Dagnino-Acosta A, Joshi AD, Knoblauch M, Xia Y, Georgiou DK, Xu J, Long C, Amano H, Reynolds C, Dong K, Martin JC, Lagor WR, Rodney GG, Sahin E, Sewry C, Hamilton SL
Nature communications 2017 Mar 24;8:14659
Nature communications 2017 Mar 24;8:14659
Calcium dysregulation, functional calpainopathy, and endoplasmic reticulum stress in sporadic inclusion body myositis.
Amici DR, Pinal-Fernandez I, Mázala DA, Lloyd TE, Corse AM, Christopher-Stine L, Mammen AL, Chin ER
Acta neuropathologica communications 2017 Mar 22;5(1):24
Acta neuropathologica communications 2017 Mar 22;5(1):24
Dysferlin mediates membrane tubulation and links T-tubule biogenesis to muscular dystrophy.
Hofhuis J, Bersch K, Büssenschütt R, Drzymalski M, Liebetanz D, Nikolaev VO, Wagner S, Maier LS, Gärtner J, Klinge L, Thoms S
Journal of cell science 2017 Mar 1;130(5):841-852
Journal of cell science 2017 Mar 1;130(5):841-852
Congenital myopathy results from misregulation of a muscle Ca2+ channel by mutant Stac3.
Linsley JW, Hsu IU, Groom L, Yarotskyy V, Lavorato M, Horstick EJ, Linsley D, Wang W, Franzini-Armstrong C, Dirksen RT, Kuwada JY
Proceedings of the National Academy of Sciences of the United States of America 2017 Jan 10;114(2):E228-E236
Proceedings of the National Academy of Sciences of the United States of America 2017 Jan 10;114(2):E228-E236
Role of calpain in eccentric contraction-induced proteolysis of Ca(2+)-regulatory proteins and force depression in rat fast-twitch skeletal muscle.
Kanzaki K, Watanabe D, Kuratani M, Yamada T, Matsunaga S, Wada M
Journal of applied physiology (Bethesda, Md. : 1985) 2017 Feb 1;122(2):396-405
Journal of applied physiology (Bethesda, Md. : 1985) 2017 Feb 1;122(2):396-405
Amphiphysin (BIN1) negatively regulates dynamin 2 for normal muscle maturation.
Cowling BS, Prokic I, Tasfaout H, Rabai A, Humbert F, Rinaldi B, Nicot AS, Kretz C, Friant S, Roux A, Laporte J
The Journal of clinical investigation 2017 Dec 1;127(12):4477-4487
The Journal of clinical investigation 2017 Dec 1;127(12):4477-4487
The SH3 and cysteine-rich domain 3 (Stac3) gene is important to growth, fiber composition, and calcium release from the sarcoplasmic reticulum in postnatal skeletal muscle.
Cong X, Doering J, Mazala DA, Chin ER, Grange RW, Jiang H
Skeletal muscle 2016;6:17
Skeletal muscle 2016;6:17
Thrombospondin expression in myofibers stabilizes muscle membranes.
Vanhoutte D, Schips TG, Kwong JQ, Davis J, Tjondrokoesoemo A, Brody MJ, Sargent MA, Kanisicak O, Yi H, Gao QQ, Rabinowitz JE, Volk T, McNally EM, Molkentin JD
eLife 2016 Sep 26;5
eLife 2016 Sep 26;5
Alternative Splicing of Four Trafficking Genes Regulates Myofiber Structure and Skeletal Muscle Physiology.
Giudice J, Loehr JA, Rodney GG, Cooper TA
Cell reports 2016 Nov 15;17(8):1923-1933
Cell reports 2016 Nov 15;17(8):1923-1933
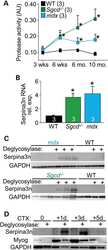
Genetic overexpression of Serpina3n attenuates muscular dystrophy in mice.
Tjondrokoesoemo A, Schips T, Kanisicak O, Sargent MA, Molkentin JD
Human molecular genetics 2016 Mar 15;25(6):1192-202
Human molecular genetics 2016 Mar 15;25(6):1192-202
Eps 15 Homology Domain (EHD)-1 Remodels Transverse Tubules in Skeletal Muscle.
Demonbreun AR, Swanson KE, Rossi AE, Deveaux HK, Earley JU, Allen MV, Arya P, Bhattacharyya S, Band H, Pytel P, McNally EM
PloS one 2015;10(9):e0136679
PloS one 2015;10(9):e0136679
Inward rectifier potassium currents in mammalian skeletal muscle fibres.
DiFranco M, Yu C, Quiñonez M, Vergara JL
The Journal of physiology 2015 Mar 1;593(5):1213-38
The Journal of physiology 2015 Mar 1;593(5):1213-38
Downregulation of connexin43 by microRNA-130a in cardiomyocytes results in cardiac arrhythmias.
Osbourne A, Calway T, Broman M, McSharry S, Earley J, Kim GH
Journal of molecular and cellular cardiology 2014 Sep;74:53-63
Journal of molecular and cellular cardiology 2014 Sep;74:53-63
EHD1 mediates vesicle trafficking required for normal muscle growth and transverse tubule development.
Posey AD Jr, Swanson KE, Alvarez MG, Krishnan S, Earley JU, Band H, Pytel P, McNally EM, Demonbreun AR
Developmental biology 2014 Mar 15;387(2):179-90
Developmental biology 2014 Mar 15;387(2):179-90
Dysferlin and myoferlin regulate transverse tubule formation and glycerol sensitivity.
Demonbreun AR, Rossi AE, Alvarez MG, Swanson KE, Deveaux HK, Earley JU, Hadhazy M, Vohra R, Walter GA, Pytel P, McNally EM
The American journal of pathology 2014 Jan;184(1):248-59
The American journal of pathology 2014 Jan;184(1):248-59
Gene therapy prolongs survival and restores function in murine and canine models of myotubular myopathy.
Childers MK, Joubert R, Poulard K, Moal C, Grange RW, Doering JA, Lawlor MW, Rider BE, Jamet T, Danièle N, Martin S, Rivière C, Soker T, Hammer C, Van Wittenberghe L, Lockard M, Guan X, Goddard M, Mitchell E, Barber J, Williams JK, Mack DL, Furth ME, Vignaud A, Masurier C, Mavilio F, Moullier P, Beggs AH, Buj-Bello A
Science translational medicine 2014 Jan 22;6(220):220ra10
Science translational medicine 2014 Jan 22;6(220):220ra10
Perturbations in intracellular Ca2+ handling in skeletal muscle in the G93A*SOD1 mouse model of amyotrophic lateral sclerosis.
Chin ER, Chen D, Bobyk KD, Mázala DA
American journal of physiology. Cell physiology 2014 Dec 1;307(11):C1031-8
American journal of physiology. Cell physiology 2014 Dec 1;307(11):C1031-8
Effects of SEA0400 on arrhythmogenicity in a Langendorff-perfused 1-month myocardial infarction rabbit model.
Chou CC, Chang PC, Wen MS, Lee HL, Chu Y, Baba A, Matsuda T, Yeh SJ, Wu D
Pacing and clinical electrophysiology : PACE 2013 May;36(5):596-606
Pacing and clinical electrophysiology : PACE 2013 May;36(5):596-606
Deregulation of the protocadherin gene FAT1 alters muscle shapes: implications for the pathogenesis of facioscapulohumeral dystrophy.
Caruso N, Herberth B, Bartoli M, Puppo F, Dumonceaux J, Zimmermann A, Denadai S, Lebossé M, Roche S, Geng L, Magdinier F, Attarian S, Bernard R, Maina F, Levy N, Helmbacher F
PLoS genetics 2013 Jun;9(6):e1003550
PLoS genetics 2013 Jun;9(6):e1003550
Dysferlin stabilizes stress-induced Ca2+ signaling in the transverse tubule membrane.
Kerr JP, Ziman AP, Mueller AL, Muriel JM, Kleinhans-Welte E, Gumerson JD, Vogel SS, Ward CW, Roche JA, Bloch RJ
Proceedings of the National Academy of Sciences of the United States of America 2013 Dec 17;110(51):20831-6
Proceedings of the National Academy of Sciences of the United States of America 2013 Dec 17;110(51):20831-6
Lack of myotubularin (MTM1) leads to muscle hypotrophy through unbalanced regulation of the autophagy and ubiquitin-proteasome pathways.
Al-Qusairi L, Prokic I, Amoasii L, Kretz C, Messaddeq N, Mandel JL, Laporte J
FASEB journal : official publication of the Federation of American Societies for Experimental Biology 2013 Aug;27(8):3384-94
FASEB journal : official publication of the Federation of American Societies for Experimental Biology 2013 Aug;27(8):3384-94
Highly conductive carbon nanotube matrix accelerates developmental chloride extrusion in central nervous system neurons by increased expression of chloride transporter KCC2.
Liedtke W, Yeo M, Zhang H, Wang Y, Gignac M, Miller S, Berglund K, Liu J
Small (Weinheim an der Bergstrasse, Germany) 2013 Apr 8;9(7):1066-75
Small (Weinheim an der Bergstrasse, Germany) 2013 Apr 8;9(7):1066-75
Domain cooperativity in the β1a subunit is essential for dihydropyridine receptor voltage sensing in skeletal muscle.
Dayal A, Bhat V, Franzini-Armstrong C, Grabner M
Proceedings of the National Academy of Sciences of the United States of America 2013 Apr 30;110(18):7488-93
Proceedings of the National Academy of Sciences of the United States of America 2013 Apr 30;110(18):7488-93
Myotubularin and PtdIns3P remodel the sarcoplasmic reticulum in muscle in vivo.
Amoasii L, Hnia K, Chicanne G, Brech A, Cowling BS, Müller MM, Schwab Y, Koebel P, Ferry A, Payrastre B, Laporte J
Journal of cell science 2013 Apr 15;126(Pt 8):1806-19
Journal of cell science 2013 Apr 15;126(Pt 8):1806-19
Abnormalities of calcium handling proteins in skeletal muscle mirror those of the heart in humans with heart failure: a shared mechanism?
Middlekauff HR, Vigna C, Verity MA, Fonarow GC, Horwich TB, Hamilton MA, Shieh P, Tupling AR
Journal of cardiac failure 2012 Sep;18(9):724-33
Journal of cardiac failure 2012 Sep;18(9):724-33
Muscle weakness in myotonic dystrophy associated with misregulated splicing and altered gating of Ca(V)1.1 calcium channel.
Tang ZZ, Yarotskyy V, Wei L, Sobczak K, Nakamori M, Eichinger K, Moxley RT, Dirksen RT, Thornton CA
Human molecular genetics 2012 Mar 15;21(6):1312-24
Human molecular genetics 2012 Mar 15;21(6):1312-24
Three-dimensional localization of the α and β subunits and of the II-III loop in the skeletal muscle L-type Ca2+ channel.
Szpyt J, Lorenzon N, Perez CF, Norris E, Allen PD, Beam KG, Samsó M
The Journal of biological chemistry 2012 Dec 21;287(52):43853-61
The Journal of biological chemistry 2012 Dec 21;287(52):43853-61
Increased expression of wild-type or a centronuclear myopathy mutant of dynamin 2 in skeletal muscle of adult mice leads to structural defects and muscle weakness.
Cowling BS, Toussaint A, Amoasii L, Koebel P, Ferry A, Davignon L, Nishino I, Mandel JL, Laporte J
The American journal of pathology 2011 May;178(5):2224-35
The American journal of pathology 2011 May;178(5):2224-35
Identification of novel ryanodine receptor 1 (RyR1) protein interaction with calcium homeostasis endoplasmic reticulum protein (CHERP).
Ryan T, Sharma P, Ignatchenko A, MacLennan DH, Kislinger T, Gramolini AO
The Journal of biological chemistry 2011 May 13;286(19):17060-8
The Journal of biological chemistry 2011 May 13;286(19):17060-8
The calcium channel α2/δ1 subunit interacts with ATP5b in the plasma membrane of developing muscle cells.
García J
American journal of physiology. Cell physiology 2011 Jul;301(1):C44-52
American journal of physiology. Cell physiology 2011 Jul;301(1):C44-52
Aquaporin-4 water channel oligomers are associated with the transverse tubules of skeletal myofibers.
Kaakinen M, Zelenin S, Metsikkö K
Experimental cell research 2011 Jan 1;317(1):20-8
Experimental cell research 2011 Jan 1;317(1):20-8
Defects in amphiphysin 2 (BIN1) and triads in several forms of centronuclear myopathies.
Toussaint A, Cowling BS, Hnia K, Mohr M, Oldfors A, Schwab Y, Yis U, Maisonobe T, Stojkovic T, Wallgren-Pettersson C, Laugel V, Echaniz-Laguna A, Mandel JL, Nishino I, Laporte J
Acta neuropathologica 2011 Feb;121(2):253-66
Acta neuropathologica 2011 Feb;121(2):253-66
Orthograde dihydropyridine receptor signal regulates ryanodine receptor passive leak.
Eltit JM, Li H, Ward CW, Molinski T, Pessah IN, Allen PD, Lopez JR
Proceedings of the National Academy of Sciences of the United States of America 2011 Apr 26;108(17):7046-51
Proceedings of the National Academy of Sciences of the United States of America 2011 Apr 26;108(17):7046-51
Aerobic exercise training improves skeletal muscle function and Ca2+ handling-related protein expression in sympathetic hyperactivity-induced heart failure.
Bueno CR Jr, Ferreira JC, Pereira MG, Bacurau AV, Brum PC
Journal of applied physiology (Bethesda, Md. : 1985) 2010 Sep;109(3):702-9
Journal of applied physiology (Bethesda, Md. : 1985) 2010 Sep;109(3):702-9
Initiating treadmill training in late middle age offers modest adaptations in Ca2+ handling but enhances oxidative damage in senescent rat skeletal muscle.
Thomas MM, Vigna C, Betik AC, Tupling AR, Hepple RT
American journal of physiology. Regulatory, integrative and comparative physiology 2010 May;298(5):R1269-78
American journal of physiology. Regulatory, integrative and comparative physiology 2010 May;298(5):R1269-78
Calsequestrin and junctin immunoreactivity in hexagonally cross-linked tubular arrays myopathy.
Di Blasi C, Blasevich F, Bellafiore E, Mottarelli E, Gibertini S, Zanotti S, Saredi S, Mantegazza R, Morandi L, Mora M
Neuromuscular disorders : NMD 2010 May;20(5):326-9
Neuromuscular disorders : NMD 2010 May;20(5):326-9
Aerobic exercise training improves Ca2+ handling and redox status of skeletal muscle in mice.
Ferreira JC, Bacurau AV, Bueno CR Jr, Cunha TC, Tanaka LY, Jardim MA, Ramires PR, Brum PC
Experimental biology and medicine (Maywood, N.J.) 2010 Apr;235(4):497-505
Experimental biology and medicine (Maywood, N.J.) 2010 Apr;235(4):497-505
Skeletal muscle type comparison of subsarcolemmal mitochondrial membrane phospholipid fatty acid composition in rat.
Stefanyk LE, Coverdale N, Roy BD, Peters SJ, LeBlanc PJ
The Journal of membrane biology 2010 Apr;234(3):207-15
The Journal of membrane biology 2010 Apr;234(3):207-15
T-tubule disorganization and defective excitation-contraction coupling in muscle fibers lacking myotubularin lipid phosphatase.
Al-Qusairi L, Weiss N, Toussaint A, Berbey C, Messaddeq N, Kretz C, Sanoudou D, Beggs AH, Allard B, Mandel JL, Laporte J, Jacquemond V, Buj-Bello A
Proceedings of the National Academy of Sciences of the United States of America 2009 Nov 3;106(44):18763-8
Proceedings of the National Academy of Sciences of the United States of America 2009 Nov 3;106(44):18763-8
Exercise-induced regulation of phospholemman (FXYD1) in rat skeletal muscle: implications for Na+/K+-ATPase activity.
Rasmussen MK, Kristensen M, Juel C
Acta physiologica (Oxford, England) 2008 Sep;194(1):67-79
Acta physiologica (Oxford, England) 2008 Sep;194(1):67-79
Reduced expression of sarcalumenin and related Ca2+ -regulatory proteins in aged rat skeletal muscle.
O'Connell K, Gannon J, Doran P, Ohlendieck K
Experimental gerontology 2008 Oct;43(10):958-61
Experimental gerontology 2008 Oct;43(10):958-61
Zebrafish relatively relaxed mutants have a ryanodine receptor defect, show slow swimming and provide a model of multi-minicore disease.
Hirata H, Watanabe T, Hatakeyama J, Sprague SM, Saint-Amant L, Nagashima A, Cui WW, Zhou W, Kuwada JY
Development (Cambridge, England) 2007 Aug;134(15):2771-81
Development (Cambridge, England) 2007 Aug;134(15):2771-81
Excitation-contraction coupling in airway smooth muscle.
Du W, McMahon TJ, Zhang ZS, Stiber JA, Meissner G, Eu JP
The Journal of biological chemistry 2006 Oct 6;281(40):30143-51
The Journal of biological chemistry 2006 Oct 6;281(40):30143-51
A Ca2+-binding domain in RyR1 that interacts with the calmodulin binding site and modulates channel activity.
Xiong L, Zhang JZ, He R, Hamilton SL
Biophysical journal 2006 Jan 1;90(1):173-82
Biophysical journal 2006 Jan 1;90(1):173-82
Neuregulin receptor ErbB2 localization at T-tubule in cardiac and skeletal muscle.
Ueda H, Oikawa A, Nakamura A, Terasawa F, Kawagishi K, Moriizumi T
The journal of histochemistry and cytochemistry : official journal of the Histochemistry Society 2005 Jan;53(1):87-91
The journal of histochemistry and cytochemistry : official journal of the Histochemistry Society 2005 Jan;53(1):87-91
Restricted distribution of mRNAs encoding a sarcoplasmic reticulum or transverse tubule protein in skeletal myofibers.
Nissinen M, Kaisto T, Salmela P, Peltonen J, Metsikkö K
The journal of histochemistry and cytochemistry : official journal of the Histochemistry Society 2005 Feb;53(2):217-27
The journal of histochemistry and cytochemistry : official journal of the Histochemistry Society 2005 Feb;53(2):217-27
Developmental induction of DHPR alpha 1s and RYR1 gene expression does not require neural or mechanical signals.
Radzyukevich TL, Cougnon MH, Moseley AE, Heiny JA
Journal of muscle research and cell motility 2004;25(1):87-94
Journal of muscle research and cell motility 2004;25(1):87-94
Regulation of dihydropyridine receptor gene expression in mouse skeletal muscles by stretch and disuse.
Radzyukevich TL, Heiny JA
American journal of physiology. Cell physiology 2004 Nov;287(5):C1445-52
American journal of physiology. Cell physiology 2004 Nov;287(5):C1445-52
Increased sensitivity of the ryanodine receptor to halothane-induced oligomerization in malignant hyperthermia-susceptible human skeletal muscle.
Glover L, Heffron JJ, Ohlendieck K
Journal of applied physiology (Bethesda, Md. : 1985) 2004 Jan;96(1):11-8
Journal of applied physiology (Bethesda, Md. : 1985) 2004 Jan;96(1):11-8
Na,K-ATPase alpha- and beta-isoform expression in developing skeletal muscles: alpha(2) correlates with t-tubule formation.
Cougnon MH, Moseley AE, Radzyukevich TL, Lingrel JB, Heiny JA
Pflugers Archiv : European journal of physiology 2002 Oct;445(1):123-31
Pflugers Archiv : European journal of physiology 2002 Oct;445(1):123-31
Homer proteins and InsP(3) receptors co-localise in the longitudinal sarcoplasmic reticulum of skeletal muscle fibres.
Salanova M, Priori G, Barone V, Intravaia E, Flucher B, Ciruela F, McIlhinney RA, Parys JB, Mikoshiba K, Sorrentino V
Cell calcium 2002 Oct;32(4):193-200
Cell calcium 2002 Oct;32(4):193-200
Homer proteins and InsP(3) receptors co-localise in the longitudinal sarcoplasmic reticulum of skeletal muscle fibres.
Salanova M, Priori G, Barone V, Intravaia E, Flucher B, Ciruela F, McIlhinney RA, Parys JB, Mikoshiba K, Sorrentino V
Cell calcium 2002 Oct;32(4):193-200
Cell calcium 2002 Oct;32(4):193-200
Sarcolemmal organization in skeletal muscle lacking desmin: evidence for cytokeratins associated with the membrane skeleton at costameres.
O'Neill A, Williams MW, Resneck WG, Milner DJ, Capetanaki Y, Bloch RJ
Molecular biology of the cell 2002 Jul;13(7):2347-59
Molecular biology of the cell 2002 Jul;13(7):2347-59
Suramin interacts with the calmodulin binding site on the ryanodine receptor, RYR1.
Papineni RV, O'Connell KM, Zhang H, Dirksen RT, Hamilton SL
The Journal of biological chemistry 2002 Dec 20;277(51):49167-74
The Journal of biological chemistry 2002 Dec 20;277(51):49167-74
Agonists cause endocytosis of nicotinic acetylcholine receptors on cultured myotubes.
St John PA, Gordon H
Journal of neurobiology 2001 Nov 15;49(3):212-23
Journal of neurobiology 2001 Nov 15;49(3):212-23
Ca2+-induced Ca2+ release supports the relay mode of activity in thalamocortical cells.
Budde T, Sieg F, Braunewell KH, Gundelfinger ED, Pape HC
Neuron 2000 May;26(2):483-92
Neuron 2000 May;26(2):483-92
Ca2+-induced Ca2+ release supports the relay mode of activity in thalamocortical cells.
Budde T, Sieg F, Braunewell KH, Gundelfinger ED, Pape HC
Neuron 2000 May;26(2):483-92
Neuron 2000 May;26(2):483-92
Extensive but coordinated reorganization of the membrane skeleton in myofibers of dystrophic (mdx) mice.
Williams MW, Bloch RJ
The Journal of cell biology 1999 Mar 22;144(6):1259-70
The Journal of cell biology 1999 Mar 22;144(6):1259-70
Cloning of a calcium channel alpha1 subunit from the reef-building coral, Stylophora pistillata.
Zoccola D, Tambutté E, Sénégas-Balas F, Michiels JF, Failla JP, Jaubert J, Allemand D
Gene 1999 Feb 18;227(2):157-67
Gene 1999 Feb 18;227(2):157-67
A transgenic myogenic cell line lacking ryanodine receptor protein for homologous expression studies: reconstitution of Ry1R protein and function.
Moore RA, Nguyen H, Galceran J, Pessah IN, Allen PD
The Journal of cell biology 1998 Feb 23;140(4):843-51
The Journal of cell biology 1998 Feb 23;140(4):843-51
A transgenic myogenic cell line lacking ryanodine receptor protein for homologous expression studies: reconstitution of Ry1R protein and function.
Moore RA, Nguyen H, Galceran J, Pessah IN, Allen PD
The Journal of cell biology 1998 Feb 23;140(4):843-51
The Journal of cell biology 1998 Feb 23;140(4):843-51
Dyspedic mouse skeletal muscle expresses major elements of the triadic junction but lacks detectable ryanodine receptor protein and function.
Buck ED, Nguyen HT, Pessah IN, Allen PD
The Journal of biological chemistry 1997 Mar 14;272(11):7360-7
The Journal of biological chemistry 1997 Mar 14;272(11):7360-7
Dyspedic mouse skeletal muscle expresses major elements of the triadic junction but lacks detectable ryanodine receptor protein and function.
Buck ED, Nguyen HT, Pessah IN, Allen PD
The Journal of biological chemistry 1997 Mar 14;272(11):7360-7
The Journal of biological chemistry 1997 Mar 14;272(11):7360-7
Branching points of renal resistance arteries are enriched in L-type calcium channels and initiate vasoconstriction.
Goligorsky MS, Colflesh D, Gordienko D, Moore LC
The American journal of physiology 1995 Feb;268(2 Pt 2):F251-7
The American journal of physiology 1995 Feb;268(2 Pt 2):F251-7
Branching points of renal resistance arteries are enriched in L-type calcium channels and initiate vasoconstriction.
Goligorsky MS, Colflesh D, Gordienko D, Moore LC
The American journal of physiology 1995 Feb;268(2 Pt 2):F251-7
The American journal of physiology 1995 Feb;268(2 Pt 2):F251-7
Alpha 1 and alpha 2 Ca2+ channel subunit expression in human neuronal and small cell carcinoma cells.
Morton ME, Cassidy TN, Froehner SC, Gilmour BP, Laurens RL
FASEB journal : official publication of the Federation of American Societies for Experimental Biology 1994 Aug;8(11):884-8
FASEB journal : official publication of the Federation of American Societies for Experimental Biology 1994 Aug;8(11):884-8
Triad formation: organization and function of the sarcoplasmic reticulum calcium release channel and triadin in normal and dysgenic muscle in vitro.
Flucher BE, Andrews SB, Fleischer S, Marks AR, Caswell A, Powell JA
The Journal of cell biology 1993 Dec;123(5):1161-74
The Journal of cell biology 1993 Dec;123(5):1161-74
Localization of the alpha 1 and alpha 2 subunits of the dihydropyridine receptor and ankyrin in skeletal muscle triads.
Flucher BE, Morton ME, Froehner SC, Daniels MP
Neuron 1990 Sep;5(3):339-51
Neuron 1990 Sep;5(3):339-51
The alpha 1 and alpha 2 polypeptides of the dihydropyridine-sensitive calcium channel differ in developmental expression and tissue distribution.
Morton ME, Froehner SC
Neuron 1989 May;2(5):1499-506
Neuron 1989 May;2(5):1499-506
Monoclonal antibody to the alpha 1-subunit of the dihydropyridine-binding complex inhibits calcium currents in BC3H1 myocytes.
Morton ME, Caffrey JM, Brown AM, Froehner SC
The Journal of biological chemistry 1988 Jan 15;263(2):613-6
The Journal of biological chemistry 1988 Jan 15;263(2):613-6
Monoclonal antibody identifies a 200-kDa subunit of the dihydropyridine-sensitive calcium channel.
Morton ME, Froehner SC
The Journal of biological chemistry 1987 Sep 5;262(25):11904-7
The Journal of biological chemistry 1987 Sep 5;262(25):11904-7
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
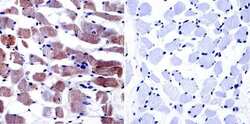
- Experimental details
- Immunohistochemistry was performed on normal biopsies of deparaffinized human skeletal muscle tissue. To expose target proteins, heat induced antigen retrieval was performed using 10mM sodium citrate (pH6.0) buffer, microwaved for 8-15 minutes. Following antigen retrieval tissues were blocked in 3% BSA-PBS for 30 minutes at room temperature. Tissues were then probed at a dilution of 1:20 with a Mouse Monoclonal Antibody recognizing Dihydropyridine Receptor alpha-1 (Product # MA3-920) or without primary antibody (negative control) overnight at 4°C in a humidified chamber. Tissues were washed extensively with PBST and endogenous peroxidase activity was quenched with a peroxidase suppressor. Detection was performed using a biotin-conjugated secondary antibody and SA-HRP, followed by colorimetric detection using DAB. Tissues were counterstained with hematoxylin and prepped for mounting.

Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
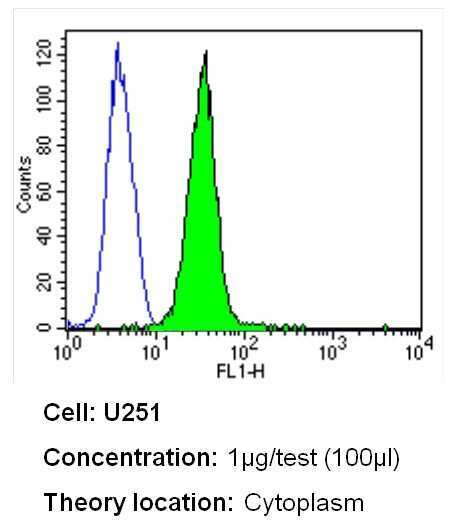
- Flow cytometry analysis of Dihydropyridine Receptor alpha-1 in U251 cells (green) compared to an isotype control (blue). Cells were harvested, adjusted to a concentration of 1-5x10^6 cells/mL, fixed with 2% paraformaldehyde and washed with PBS. Cells were blocked with a 2% solution of BSA-PBS for 30 min at room temperature and incubated with a Dihydropyridine Receptor alpha-1 monoclonal antibody (Product # MA3-920) at a dilution of 1 µg/test for 40 min at room temperature. Cells were then incubated for 40 min at room temperature in the dark using a Dylight 488-conjugated secondary antibody and re-suspended in PBS for FACS analysis.
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
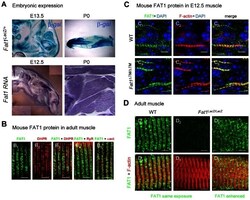
- Figure 7 Fat1 expression at late stages of muscle differentiation. ( A ) Fat1 expression was visualized in E13.5 embryos or in neonate (P0) muscle by beta-galactosidase staining or by in situ hybridization with a Fat1 3'UTR RNA probe. ( B - D ) Immunolocalization of FAT1 (anti-FAT1-ICD from [35] , green) was performed in E12.5 mouse embryo ( C ), and on adult ( B, D ) muscle fibers on longitudinal muscle cryosections from wild type ( B , C 1-3 , D 1,4 ), from Fat1 DeltaTM/DeltaTM embryos ( C 4-6 ), and from Fat1 LacZ/LacZ ( D 2-3 , D 5-6 ) mice, combined with either antibodies against alpha-actinin (red, B 5 ), DHPR (Cacna1s) (red, B 2,3 ), or RyR (red, B 4 ), or with Phalloidin (red, C, D ). In D , Green channel images (FAT1) were first captured with either identical exposure time between wild type and mutants ( D 1,4 and D 2,5 , 421 ms), or with longer exposure time ( D 3,6 , 2222 ms). This indicates that the epitope detected by the anti-FAT1-ICD antibody (from ref [35] ) is present in reduced but detectable amounts in Fat1 LacZ/LacZ muscles. This observation was made when Fat1 LacZ/LacZ mice (n = 2 at P0; and n = 3 at adult stages) displayed severe muscle defects at the stage of dissection, indicating that levels of FAT1 protein inversely correlate with phenotype severity. Scale bars: ( B - D ) 4 um, ( C ) 6 um.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
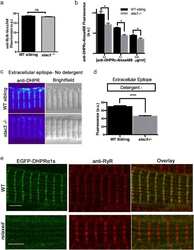
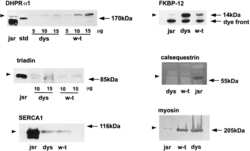
- Fig 2 Misexpression of triad proteins and expansion of the T-tubule compartment in Ehd1- heterozygous muscle. (A) BIN1 and DHPR levels are increased +2.25 and +5.4 fold in Ehd1 +/- quadriceps muscle compared to WT correlating with the increase in T-tubule structures. Gel code bands are shown as a loading control (LC). (B) Junctophilin 2 (JP2) protein levels were decreased 13-fold in Ehd1+/- quadriceps muscle compared to wildtype controls. Gel code stained bands are shown as a loading control (LC). (C) Junctophilin 1 (JP1) protein levels were similar in Ehd1+/- quadriceps muscle compared to wildtype controls. Gel code stained bands are shown as a loading control (LC). (D) Ultrastructural analysis reveals ectopic (dotted arrow) and elongated (arrow) T-tubules in 8-week-old Ehd1- heterozygous muscle ( Ehd1+/- ) stained with potassium ferricyanide to color the T-tubule structures black. (E) Ehd1- heterozygous muscle contains duplicated triads containing 2 T-tubules (black arrows) and 3 sarcoplasmic reticulum (SR) in 1 triad unit. Scale 0.5mum. (F) Ehd1- heterozygous muscle stained with potassium ferricyanide, outlines duplicated T-tubule structures (two black arrows). Scale 0.5mum. (G) Ultrastructural analysis of 2-D images reveals increased tubule abnormalities in Ehd1- heterozygous muscle, 12.5%, compared to 1.7% in control muscle (n>400 structures per genotype, p = 0.04).

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
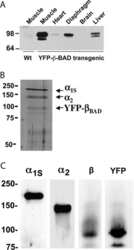
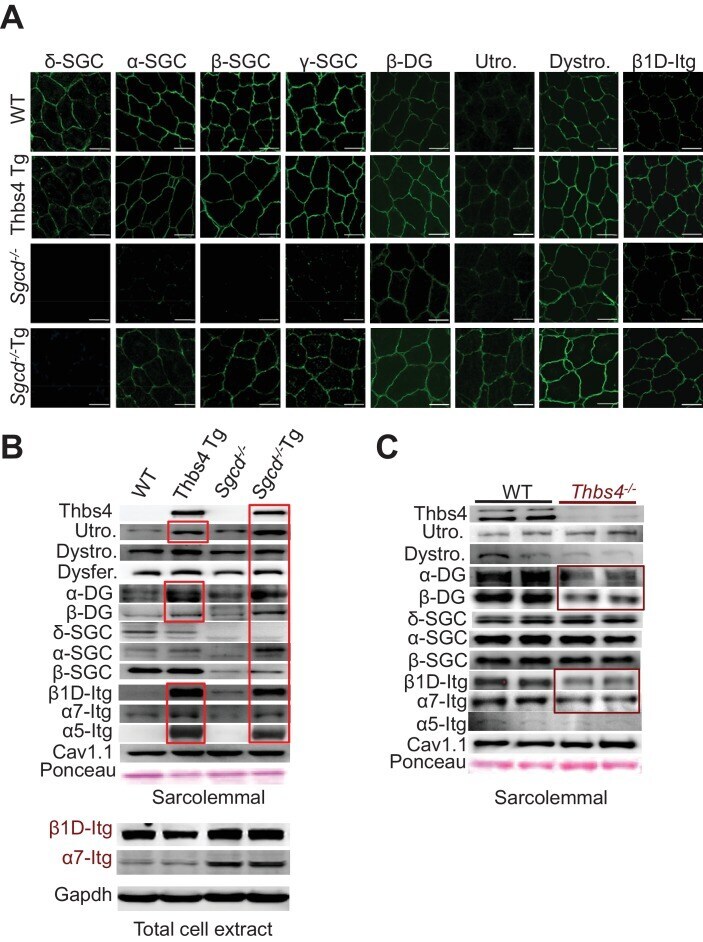
- Figure 6. Thbs4 enhances stabilizing proteins at the sarcolemma. ( A ) Immunofluoresence (green) detection of delta-, alpha-, beta-, and gamma-sarcoclycan (SGC), beta-dystroglycan (beta-DG), utrophin (Utro.), dystrophin (Dystro.) and beta1D-integrin in littermates of three month-old WT, Thbs4-Tg, Sgcd -/- and Sgcd -/- Thbs4-Tg quadriceps. Representative images of 4 mice per genotype are shown. Scale bar = 25 mum. ( B ) Representative Western blots of sarcolemmal protein extracts (upper) or total cytoplasmic protein extracts (lower) from the quadriceps of the indicated groups of mice for the indicated proteins (n = 4-5 biological replicates). Abbreviations: Utro, utrophin; Dystro, dystrophin; Dysfer, dysferlin; alpha-DG, alpha-dystroglycan; beta-DG, beta-dystroglycan; delta-SCG, delta-sarcoglycan; alpha-SCG, alpha-sarcoglycan; beta-SGC, beta-sarcoglycan; beta1D-, alpha7- and alpha5- integrin. The red boxes show increased protein levels. Also see Figure 6--figure supplement 1 for replicates. ( C ) Representative immunoblotting for structural components of the DGC- and integrin-associated protein complexes in sarcolemmal preparations from Thbs4 -/- and WT quadriceps at four months of age (n = 4 biological replicates). The burgundy-boxed areas show reduced protein levels. Ponceau staining of a nonspecific band and dihydropyridine receptor alpha1 (Cav1.1) were used as loading controls for sarcolemmal protein extracts; Gapdh was used as loading control for total cell protein extrac
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
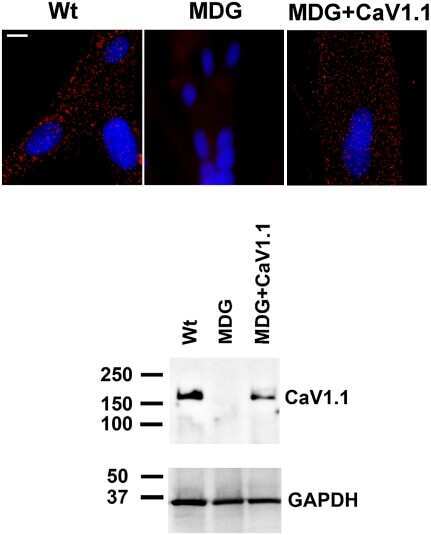
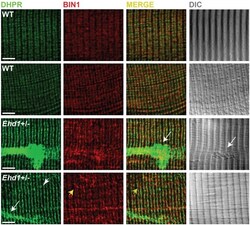
- Fig 1 Disordered T-tubules in Ehd1- heterozygous muscle. Myofibers were immunostained with anti-BIN1 (red) and anti-DHPR (green) antibodies. Representative myofibers are shown. Ehd1- heterozygous ( Ehd1+/- ) fibers displayed disorganized (white arrowhead) and aggregated (white arrow) T-tubule structures in 27% of myofibers, marked by DHPR, also evidenced in DIC images compared to 0% in control fibers. Ehd1 -heterozygous muscle with extensive BIN1 fluorescence extending beyond DHPR staining (yellow arrowhead). Scale 5mum.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
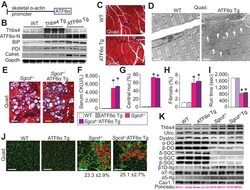
- Figure 7. Skeletal muscle specific ATF6alpha overexpression drives ER stress and intracellular vesicular expansion, but not protection against MD. ( A ) Schematic diagram of the transgene (Tg) used to overexpress ATF6alpha in skeletal muscle. ( B ) Western blot analysis for Thbs4, ATF6alpha, BiP, PDI and calreticulin (Calret.) expression in quadriceps (Quad) from WT, Thbs4 Tg and ATF6alpha Tg mice at six weeks of age. Gapdh is a processing and loading control (n = 6 biological replicates). ( C ) Masson's trichrome-stained histological sections from Quad of WT and ATF6alpha-Tg littermates at six weeks of age. Representative images of 5 mice per genotype are shown. Scale bar = 100 mum. ( D ) Transmission electron micrographs in Quad from WT and ATF6alpha-Tg mice at six weeks of age. The white arrows show dramatic expansion of ER and associated vesicles throughout the cell and especially in the sub-sarcolemmal region. Representative images of 2 mice per genotype are shown. Scale bar = 2 mum. ( E ) Masson's trichrome stained histological sections of Quad from Sgcd -/- and Sgcd -/- ATF6alpha Tg mice at six weeks of age. Representative images of 5 mice per genotype are shown. Scale bar = 100 mum. ( F ) Quantitation of serum CK levels (units/liter) in the indicated genotypes of mice shown in the legend below the graph at six weeks of age. n = 10 mice for WT and n = 8 mice for the remaining genotypes. *p

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
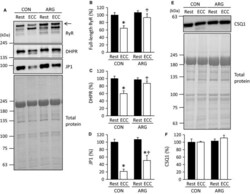
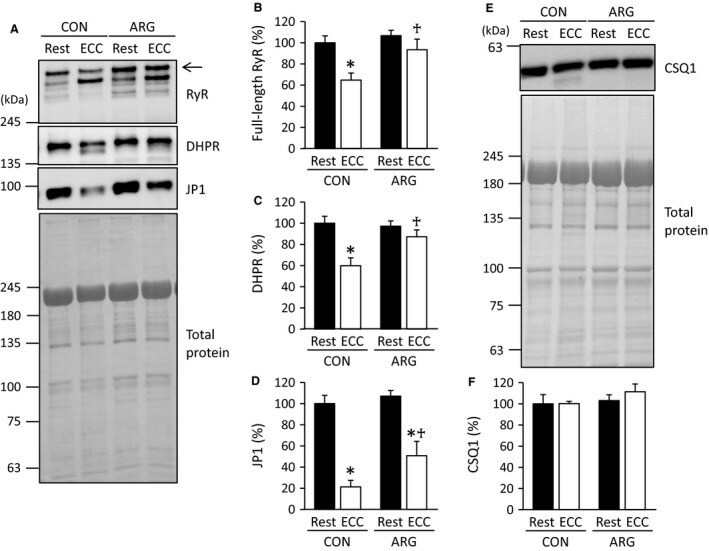
- Figure 2 ARG ingestion inhibits ECC -induced proteolysis of Ca 2+ regulatory proteins. For the protocols of ARG ingestion and ECC , see legend of Figure 1 . (A and E) immunoblot analyses of RyR, DHPR , JP 1, and CSQ 1 in tibialis anterior muscles. RyR, DHPR , and JP 1 were analyzed on the same membrane. For RyR and DHPR , in addition to full-length proteins, faster migrating bands were observed and most likely corresponded to degraded proteins. Total proteins on the membrane, which were used as a loading control, were visualized with Coomassie blue R staining. Positions of molecular mass makers are indicated on the left. (B, C, D, and F) means +- SEM ( n = 7 for each muscle) of the contents of full-length RyR (arrow), DHPR , JP 1, and CSQ 1, respectively. The contents of four proteins investigated were evaluated relative to total proteins. The results are expressed as percentages of the values observed in rested muscles from CON rats. * P < 0.05, versus rested muscles within rats; + P < 0.05, versus ECC muscles from CON rats. ARG , l -arginine; CON , control; CSQ 1, calsequestrin-1; DHPR , dihydropyridine receptor; ECC , eccentric contraction; JP 1, junctophilin-1; RyR, ryanodine receptor.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- Figure 8 In vitro S-nitrosylation of calpain-1 inhibits Ca2+-induced proteolysis of Ca2+ regulatory proteins. (A) immunodetection of GSNO-induced S-nitrosylation of calpain-1 in tibialis anterior muscles. To check the assay specificity and ascertain whether GSNO treatment can result in S-nitrosylation of calpain-1, SNO-RAC was performed in the presence (+) and absence (−) of GSNO and Cu-Asc treatment. Positions of molecular mass markers are indicated on the left. (B) immunoblots of RyR, DHPR, and JP1. Muscle homogenates were incubated in the presence (+) and absence (−) of GSNO and Ca2+. Total proteins on the membrane, which were used as a loading control, were visualized with Coomassie blue R staining. (C, D and E) means ± SEM (n = 5 for each treatment) of the contents of full-length RyR (arrow), DHPR, and JP1, respectively. The contents of the three proteins were evaluated relative to the total proteins. The results are expressed as percentages of the values in samples treated without GSNO and Ca2+. a P < 0.05, versus T1. b P < 0.05, versus T2. c P < 0.05, versus T3. GSNO, S-nitrosoglutathione; Cu-Asc, CuCl and sodium ascorbate; DHPR, dihydropyridine receptor; JP1, junctophilin-1; RyR, ryanodine receptor; SNO-RAC, resin-assisted capture of S-nitrosothiols; T, treatment.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
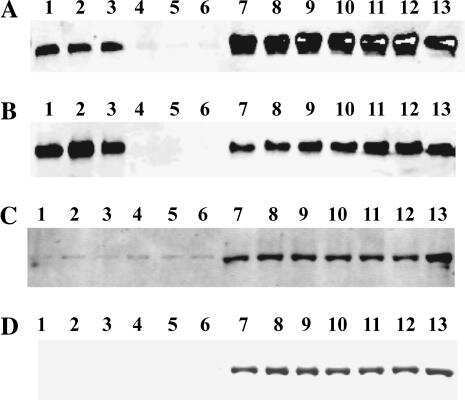
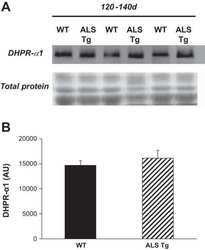
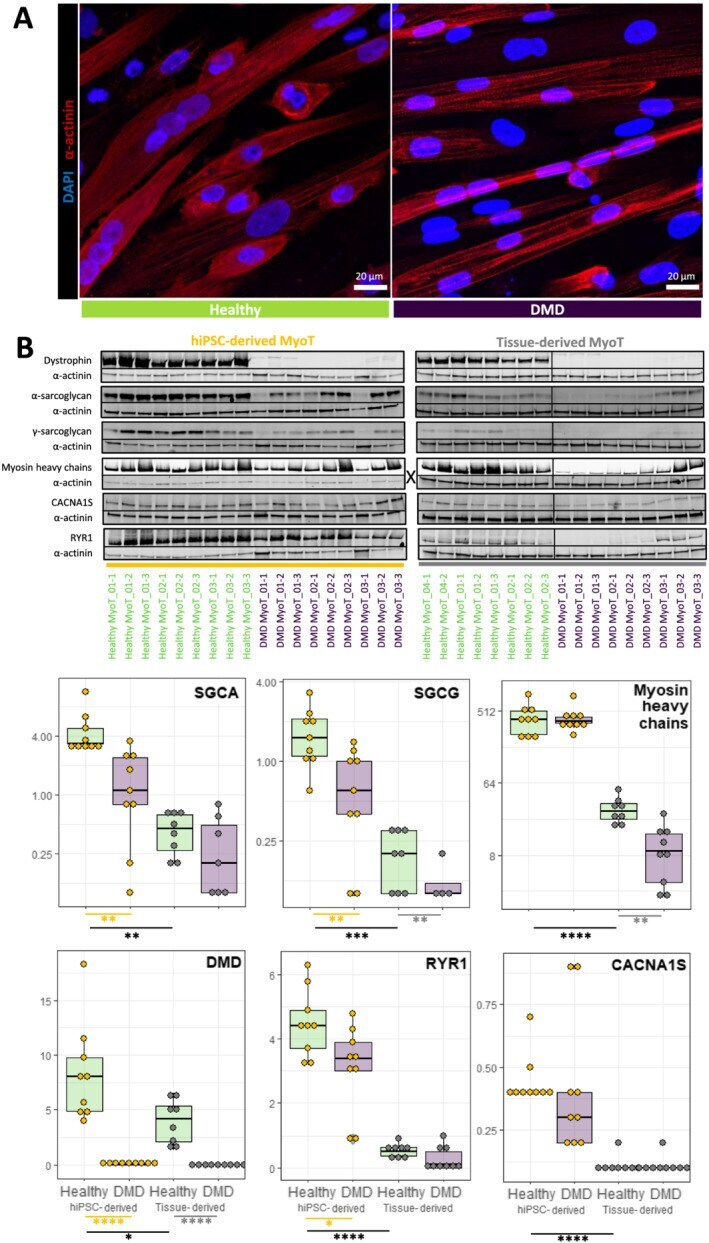
- Figure 3 Comparison of healthy and Duchenne muscular dystrophy (DMD) MyoT from hiPSCs and tissues at the protein level. (A) hiPSC-derived MyoT immunolabelling of alpha-actinin (red) and nuclei (DAPI, blue) in healthy (left) and DMD cells (right). (B) Representative western blots and related quantifications of DMD, SGCA, SGCG, myosin heavy chains, CACNA1S, and RYR1 from protein extracts in healthy and DMD hiPSC-derived and tissue-derived MyoT (X: 0.25 mug of total protein was used in hiPSC-derived MyoT instead of 7 mug in tissue-derived MyoT--* P value
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
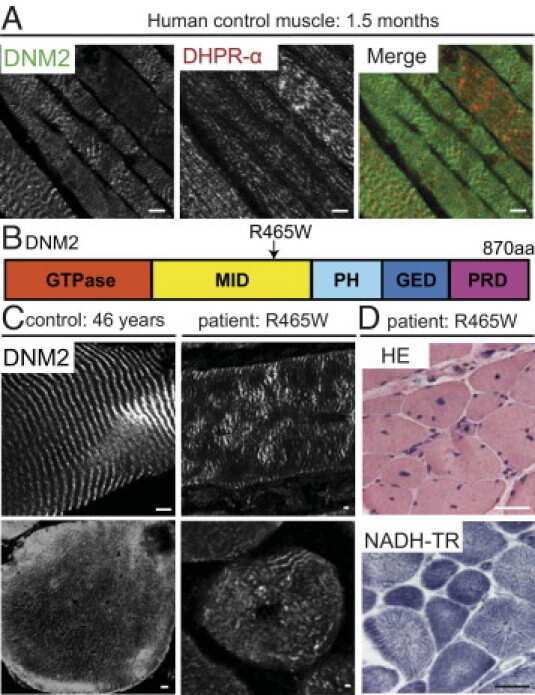
- Figure 9 FAT1 protein and RNA levels are mis-regulated in human foetal FSHD tissues. ( A ) Immunolocalization of FAT1 (Rb-1465 anti FAT1-ICD, green) and DHPR (Cacna1s, magenta) in longitudinal sections from human quadriceps biopsies from a control (top) or and FSHD (F1, bottom) foetus with 1.5 D4Z4 repeats. ( B ) qPCR analysis of FAT1 mRNA levels in quadriceps (3 left graphs) and deltoid muscles (middle graph) and in brain (right graph), comparing respectively with age-matched control foetuses (blue bars), a 26 weeks old FSHD1 foetus (F1) harbouring 1.5 D4Z4 repeats in the 4q35 region (dark red bars), a 16 weeks old FSHD1 foetus harbouring 4.3 D4Z4 repeats at 4q35 region (F2), and twin FSHD1 foetuses aged 28 weeks, with 7 D4Z4 repeats. ( C ) Analysis of the regulatory status of the promoter region by Chromatin immunoprecipitation. The respective level of the following histone marks: H3K27me3 (silenced chromatin; C-left ), and H3K4m3 (promoter active; C-right ), in muscle extracts from four age matched controls (ct1 to 4) or four FSHD1 foetuses (F1 to F4) are shown. Relative quantities were normalized with the level of histone marks at the promoter of the GUSB gene as internal control, and expressed as % of control 1 (ct1). Scale bars: ( A ) 50 um.

- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
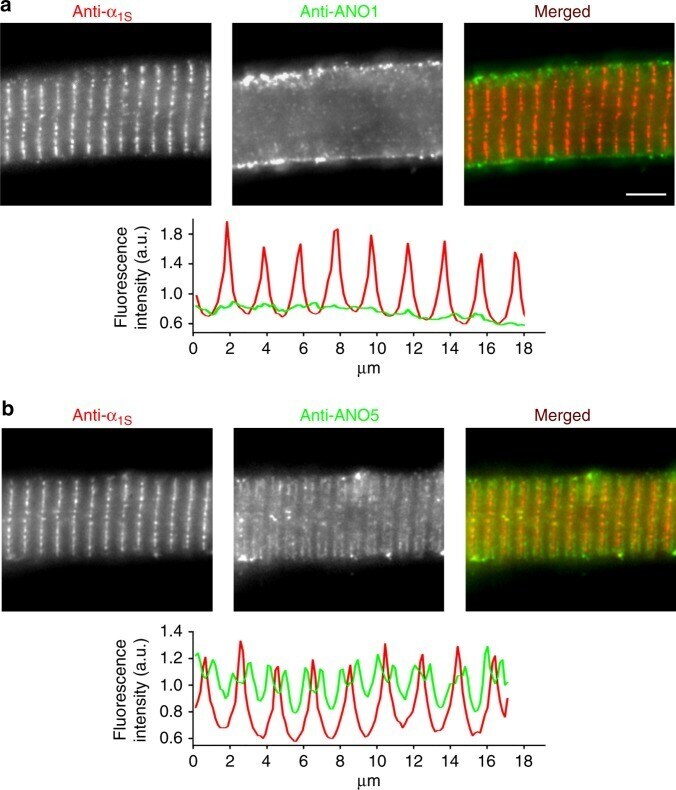
- Fig. 5 ANO1 is localised in the zebrafish skeletal muscle surface membrane. Representative images of normal zebrafish myotubes double immunolabelled for DHPRalpha 1S subunit (anti-alpha 1S ) and ANO1 (anti-ANO1) or ANO5 (anti-ANO5) isoforms showing their respective subcellular distribution. a ANO1 is expressed in the sarcolemma (centre image) and does not co-localise with DHPRalpha 1S (left image) in the triads (merged image). Scale bar, 5 um. Fluorescence intensity profile (bottom) obtained by measuring the average fluorescence intensity along a horizontal line on the merged image demonstrates regular t-tubular periodicity of the DHPRalpha 1S (red) but a complete lack of a structured ANO1 signal (green). b Double immunofluorescence labelling of DHPRalpha 1S (left image) and ANO5 (centre image) under similar conditions clearly revealed anti-ANO5 signal on both sides of the t-tubular membrane (merged image) and thus indicating SR membrane localisation of ANO5. Fluorescence intensity profile displays distinct but regular spatial periodicity with one red peak (DHPR signal) flanked by two green peaks (ANO5 signal)