Explore
Explore Validate
Validate Learn
Learn Western blot
Western blotAntibody data
- Antibody Data
- Antigen structure
- References [30]
- Comments [0]
- Validations
- Western blot [1]
- Immunocytochemistry [3]
- Other assay [18]
Submit
Validation data
Reference
Comment
Report error
- Product number
- 14-6503-80 - Provider product page

- Provider
- Invitrogen Antibodies
- Product name
- Myosin 4 Monoclonal Antibody (MF20), eBioscience™
- Antibody type
- Monoclonal
- Antigen
- Other
- Reactivity
- Human, Mouse, Rat, Bovine, Canine, Chicken/Avian, Feline, Guinea Pig, Rabbit
- Host
- Mouse
- Isotype
- IgG
- Antibody clone number
- MF20
- Vial size
- 25 µg
- Concentration
- 0.5 mg/mL
- Storage
- 4° C
Submitted references Severe congenital myasthenic syndromes caused by agrin mutations affecting secretion by motoneurons.
Spatiotemporal single-cell RNA sequencing of developing chicken hearts identifies interplay between cellular differentiation and morphogenesis.
TAM kinase signaling is indispensable for proper skeletal muscle regeneration in mice.
Hydrostatic mechanical stress regulates growth and maturation of the atrioventricular valve.
hiPSC-Derived Neurons Provide a Robust and Physiologically Relevant In Vitro Platform to Test Botulinum Neurotoxins.
Possible application of muscle specific conditional mouse-derived induced pluripotent stem cells for muscle research.
TAK-242, a specific inhibitor of Toll-like receptor 4 signalling, prevents endotoxemia-induced skeletal muscle wasting in mice.
Stable Knockdown of Genes Encoding Extracellular Matrix Proteins in the C2C12 Myoblast Cell Line Using Small-Hairpin (sh)RNA.
Loss of ARNT in skeletal muscle limits muscle regeneration in aging.
Data on the proliferation and differentiation of C2C12 myoblast treated with branched-chain ketoacid dehydrogenase kinase inhibitor.
In vivo detection of programmed cell death during mouse heart development.
Bmi1 inhibitor PTC-209 promotes Chemically-induced Direct Cardiac Reprogramming of cardiac fibroblasts into cardiomyocytes.
Sema3a-Nrp1 Signaling Mediates Fast-Twitch Myofiber Specificity of Tw2(+) Cells.
Genome-wide microhomologies enable precise template-free editing of biologically relevant deletion mutations.
IRE1-XBP1 Pathway of the Unfolded Protein Response Is Required during Early Differentiation of C2C12 Myoblasts.
Cell surface flip-flop of phosphatidylserine is critical for PIEZO1-mediated myotube formation.
Peptidyl-prolyl cis-trans isomerase NIMA interacting 1 regulates skeletal muscle fusion through structural modification of Smad3 in the linker region.
Autologous Cell Therapy Approach for Duchenne Muscular Dystrophy using PiggyBac Transposons and Mesoangioblasts.
Lipopolysaccharide inhibits myogenic differentiation of C2C12 myoblasts through the Toll-like receptor 4-nuclear factor-κB signaling pathway and myoblast-derived tumor necrosis factor-α.
The requirement of Mettl3-promoted MyoD mRNA maintenance in proliferative myoblasts for skeletal muscle differentiation.
FTO is required for myogenesis by positively regulating mTOR-PGC-1α pathway-mediated mitochondria biogenesis.
Proteolysis regulates cardiomyocyte maturation and tissue integration.
Ultrastructure of medial rectus muscles in patients with intermittent exotropia.
Single-molecule analysis of myocyte differentiation reveals bimodal lineage commitment.
Inhibition of TGFβ signaling increases direct conversion of fibroblasts to induced cardiomyocytes.
iPS cell-derived cardiogenicity is hindered by sustained integration of reprogramming transgenes.
Optimization of direct fibroblast reprogramming to cardiomyocytes using calcium activity as a functional measure of success.
Myocardin is a master regulator of smooth muscle gene expression.
Myosins: a diverse superfamily.
Immunochemical analysis of myosin heavy chain during avian myogenesis in vivo and in vitro.
Jacquier A, Risson V, Simonet T, Roussange F, Lacoste N, Ribault S, Carras J, Theuriet J, Girard E, Grosjean I, Le Goff L, Kröger S, Meltoranta J, Bauché S, Sternberg D, Fournier E, Kostera-Pruszczyk A, O'Connor E, Eymard B, Lochmüller H, Martinat C, Schaeffer L
Acta neuropathologica 2022 Oct;144(4):707-731
Acta neuropathologica 2022 Oct;144(4):707-731
Spatiotemporal single-cell RNA sequencing of developing chicken hearts identifies interplay between cellular differentiation and morphogenesis.
Mantri M, Scuderi GJ, Abedini-Nassab R, Wang MFZ, McKellar D, Shi H, Grodner B, Butcher JT, De Vlaminck I
Nature communications 2021 Mar 19;12(1):1771
Nature communications 2021 Mar 19;12(1):1771
TAM kinase signaling is indispensable for proper skeletal muscle regeneration in mice.
Al-Zaeed N, Budai Z, Szondy Z, Sarang Z
Cell death & disease 2021 Jun 12;12(6):611
Cell death & disease 2021 Jun 12;12(6):611
Hydrostatic mechanical stress regulates growth and maturation of the atrioventricular valve.
Bassen D, Wang M, Pham D, Sun S, Rao R, Singh R, Butcher J
Development (Cambridge, England) 2021 Jul 1;148(13)
Development (Cambridge, England) 2021 Jul 1;148(13)
hiPSC-Derived Neurons Provide a Robust and Physiologically Relevant In Vitro Platform to Test Botulinum Neurotoxins.
Lamotte JD, Roqueviere S, Gautier H, Raban E, Bouré C, Fonfria E, Krupp J, Nicoleau C
Frontiers in pharmacology 2020;11:617867
Frontiers in pharmacology 2020;11:617867
Possible application of muscle specific conditional mouse-derived induced pluripotent stem cells for muscle research.
Hosoyama T
Biochemistry and biophysics reports 2020 Mar;21:100744
Biochemistry and biophysics reports 2020 Mar;21:100744
TAK-242, a specific inhibitor of Toll-like receptor 4 signalling, prevents endotoxemia-induced skeletal muscle wasting in mice.
Ono Y, Maejima Y, Saito M, Sakamoto K, Horita S, Shimomura K, Inoue S, Kotani J
Scientific reports 2020 Jan 20;10(1):694
Scientific reports 2020 Jan 20;10(1):694
Stable Knockdown of Genes Encoding Extracellular Matrix Proteins in the C2C12 Myoblast Cell Line Using Small-Hairpin (sh)RNA.
Taye N, Stanley S, Hubmacher D
Journal of visualized experiments : JoVE 2020 Feb 12;(156)
Journal of visualized experiments : JoVE 2020 Feb 12;(156)
Loss of ARNT in skeletal muscle limits muscle regeneration in aging.
Endo Y, Baldino K, Li B, Zhang Y, Sakthivel D, MacArthur M, Panayi AC, Kip P, Spencer DJ, Jasuja R, Bagchi D, Bhasin S, Nuutila K, Neppl RL, Wagers AJ, Sinha I
FASEB journal : official publication of the Federation of American Societies for Experimental Biology 2020 Dec;34(12):16086-16104
FASEB journal : official publication of the Federation of American Societies for Experimental Biology 2020 Dec;34(12):16086-16104
Data on the proliferation and differentiation of C2C12 myoblast treated with branched-chain ketoacid dehydrogenase kinase inhibitor.
Sato Y, Tate H, Yoshizawa F, Sato Y
Data in brief 2020 Aug;31:105766
Data in brief 2020 Aug;31:105766
In vivo detection of programmed cell death during mouse heart development.
Martínez-Lagunas K, Yamaguchi Y, Becker C, Geisen C, DeRuiter MC, Miura M, Fleischmann BK, Hesse M
Cell death and differentiation 2020 Apr;27(4):1398-1414
Cell death and differentiation 2020 Apr;27(4):1398-1414
Bmi1 inhibitor PTC-209 promotes Chemically-induced Direct Cardiac Reprogramming of cardiac fibroblasts into cardiomyocytes.
Testa G, Russo M, Di Benedetto G, Barbato M, Parisi S, Pirozzi F, Tocchetti CG, Abete P, Bonaduce D, Russo T, Passaro F
Scientific reports 2020 Apr 28;10(1):7129
Scientific reports 2020 Apr 28;10(1):7129
Sema3a-Nrp1 Signaling Mediates Fast-Twitch Myofiber Specificity of Tw2(+) Cells.
Li S, Karri D, Sanchez-Ortiz E, Jaichander P, Bassel-Duby R, Liu N, Olson EN
Developmental cell 2019 Oct 7;51(1):89-98.e4
Developmental cell 2019 Oct 7;51(1):89-98.e4
Genome-wide microhomologies enable precise template-free editing of biologically relevant deletion mutations.
Grajcarek J, Monlong J, Nishinaka-Arai Y, Nakamura M, Nagai M, Matsuo S, Lougheed D, Sakurai H, Saito MK, Bourque G, Woltjen K
Nature communications 2019 Oct 24;10(1):4856
Nature communications 2019 Oct 24;10(1):4856
IRE1-XBP1 Pathway of the Unfolded Protein Response Is Required during Early Differentiation of C2C12 Myoblasts.
Tokutake Y, Yamada K, Hayashi S, Arai W, Watanabe T, Yonekura S
International journal of molecular sciences 2019 Dec 26;21(1)
International journal of molecular sciences 2019 Dec 26;21(1)
Cell surface flip-flop of phosphatidylserine is critical for PIEZO1-mediated myotube formation.
Tsuchiya M, Hara Y, Okuda M, Itoh K, Nishioka R, Shiomi A, Nagao K, Mori M, Mori Y, Ikenouchi J, Suzuki R, Tanaka M, Ohwada T, Aoki J, Kanagawa M, Toda T, Nagata Y, Matsuda R, Takayama Y, Tominaga M, Umeda M
Nature communications 2018 May 24;9(1):2049
Nature communications 2018 May 24;9(1):2049
Peptidyl-prolyl cis-trans isomerase NIMA interacting 1 regulates skeletal muscle fusion through structural modification of Smad3 in the linker region.
Islam R, Yoon H, Shin HR, Bae HS, Kim BS, Yoon WJ, Woo KM, Baek JH, Lee YS, Ryoo HM
Journal of cellular physiology 2018 Dec;233(12):9390-9403
Journal of cellular physiology 2018 Dec;233(12):9390-9403
Autologous Cell Therapy Approach for Duchenne Muscular Dystrophy using PiggyBac Transposons and Mesoangioblasts.
Iyer PS, Mavoungou LO, Ronzoni F, Zemla J, Schmid-Siegert E, Antonini S, Neff LA, Dorchies OM, Jaconi M, Lekka M, Messina G, Mermod N
Molecular therapy : the journal of the American Society of Gene Therapy 2018 Apr 4;26(4):1093-1108
Molecular therapy : the journal of the American Society of Gene Therapy 2018 Apr 4;26(4):1093-1108
Lipopolysaccharide inhibits myogenic differentiation of C2C12 myoblasts through the Toll-like receptor 4-nuclear factor-κB signaling pathway and myoblast-derived tumor necrosis factor-α.
Ono Y, Sakamoto K
PloS one 2017;12(7):e0182040
PloS one 2017;12(7):e0182040
The requirement of Mettl3-promoted MyoD mRNA maintenance in proliferative myoblasts for skeletal muscle differentiation.
Kudou K, Komatsu T, Nogami J, Maehara K, Harada A, Saeki H, Oki E, Maehara Y, Ohkawa Y
Open biology 2017 Sep;7(9)
Open biology 2017 Sep;7(9)
FTO is required for myogenesis by positively regulating mTOR-PGC-1α pathway-mediated mitochondria biogenesis.
Wang X, Huang N, Yang M, Wei D, Tai H, Han X, Gong H, Zhou J, Qin J, Wei X, Chen H, Fang T, Xiao H
Cell death & disease 2017 Mar 23;8(3):e2702
Cell death & disease 2017 Mar 23;8(3):e2702
Proteolysis regulates cardiomyocyte maturation and tissue integration.
Fukuda R, Gunawan F, Beisaw A, Jimenez-Amilburu V, Maischein HM, Kostin S, Kawakami K, Stainier DY
Nature communications 2017 Feb 17;8:14495
Nature communications 2017 Feb 17;8:14495
Ultrastructure of medial rectus muscles in patients with intermittent exotropia.
Yao J, Wang X, Ren H, Liu G, Lu P
Eye (London, England) 2016 Jan;30(1):146-51
Eye (London, England) 2016 Jan;30(1):146-51
Single-molecule analysis of myocyte differentiation reveals bimodal lineage commitment.
Gibson TM, Gersbach CA
Integrative biology : quantitative biosciences from nano to macro 2015 Jun;7(6):663-71
Integrative biology : quantitative biosciences from nano to macro 2015 Jun;7(6):663-71
Inhibition of TGFβ signaling increases direct conversion of fibroblasts to induced cardiomyocytes.
Ifkovits JL, Addis RC, Epstein JA, Gearhart JD
PloS one 2014;9(2):e89678
PloS one 2014;9(2):e89678
iPS cell-derived cardiogenicity is hindered by sustained integration of reprogramming transgenes.
Martinez-Fernandez A, Nelson TJ, Reyes S, Alekseev AE, Secreto F, Perez-Terzic C, Beraldi R, Sung HK, Nagy A, Terzic A
Circulation. Cardiovascular genetics 2014 Oct;7(5):667-76
Circulation. Cardiovascular genetics 2014 Oct;7(5):667-76
Optimization of direct fibroblast reprogramming to cardiomyocytes using calcium activity as a functional measure of success.
Addis RC, Ifkovits JL, Pinto F, Kellam LD, Esteso P, Rentschler S, Christoforou N, Epstein JA, Gearhart JD
Journal of molecular and cellular cardiology 2013 Jul;60:97-106
Journal of molecular and cellular cardiology 2013 Jul;60:97-106
Myocardin is a master regulator of smooth muscle gene expression.
Wang Z, Wang DZ, Pipes GC, Olson EN
Proceedings of the National Academy of Sciences of the United States of America 2003 Jun 10;100(12):7129-34
Proceedings of the National Academy of Sciences of the United States of America 2003 Jun 10;100(12):7129-34
Myosins: a diverse superfamily.
Sellers JR
Biochimica et biophysica acta 2000 Mar 17;1496(1):3-22
Biochimica et biophysica acta 2000 Mar 17;1496(1):3-22
Immunochemical analysis of myosin heavy chain during avian myogenesis in vivo and in vitro.
Bader D, Masaki T, Fischman DA
The Journal of cell biology 1982 Dec;95(3):763-70
The Journal of cell biology 1982 Dec;95(3):763-70
No comments: Submit comment
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
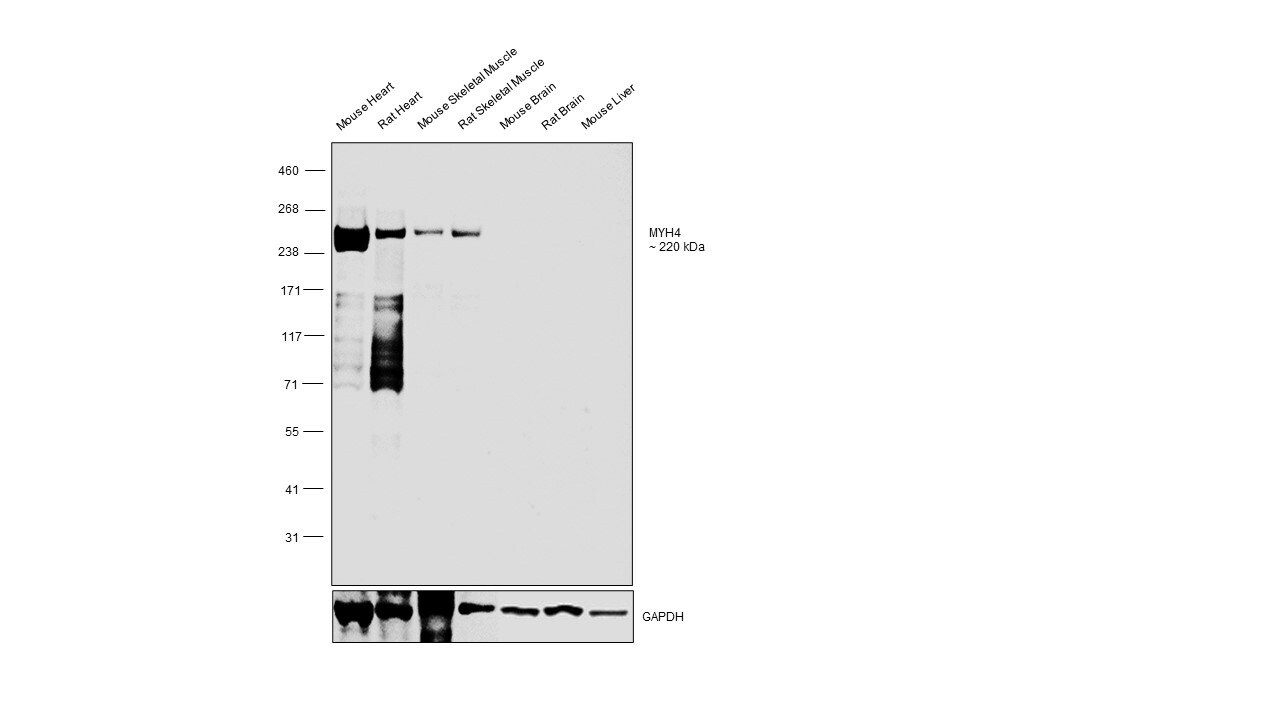
- Western blot was performed using Myosin 4 Monoclonal Antibody (MF20), eBioscience™ (Product # 14-6503-82) and a ~230 kDa band corresponding to MYH4 was observed across tissues tested except Mouse Brain, Rat Brain and Mouse Liver which are reported negative. Whole cell extracts (30 µg lysate) of Mouse Heart (Lane 1), Rat Heart (Lane 2), Mouse Skeletal Muscle (Lane 3), Rat Skeletal Muscle (Lane 4), Mouse Brain (Lane 5), Rat Brain (Lane 6), Mouse Liver (Lane 7) were electrophoresed using NuPAGE™ 3 to 8%, Tris-Acetate, 1.0–1.5 mm, Mini Protein Gels (Product # EA03752BOX), 12 well. Resolved proteins were then transferred onto a nitrocellulose membrane (Product # IB23001) by iBlot® 2 Dry Blotting System (Product # IB21001). The blot was probed with the primary antibody (1:1000 dilution) and detected by chemiluminescence with Goat anti-Mouse IgG (H+L) Superclonal™ Recombinant Secondary Antibody, HRP (Product # A28177, 1:20,000 dilution) using the iBright™ FL1500 Imaging System (Product # A44115). Chemiluminescent detection was performed using SuperSignal™ West Pico PLUS Chemiluminescent Substrate (Product # 34580).
Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
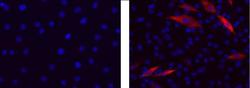
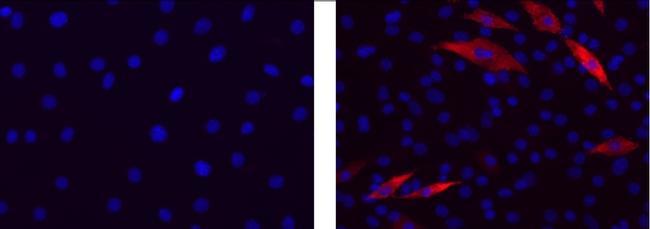
- Immunocytochemistry of differentiated C2C12 cells using 1 µg/mL of Mouse IgG2b K Isotype Control (left) or 1 µg/mL Anti-Myosin Heavy Chain Purified (right) followed by Anti-Mouse IgG TRITC.Nuclei are stained with DAPI.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
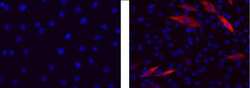
- Experimental details
- Immunocytochemistry of differentiated C2C12 cells using 1 µg/mL of Mouse IgG2b K Isotype Control (left) or 1 µg/mL Anti-Myosin Heavy Chain Purified (right) followed by Anti-Mouse IgG TRITC.Nuclei are stained with DAPI.
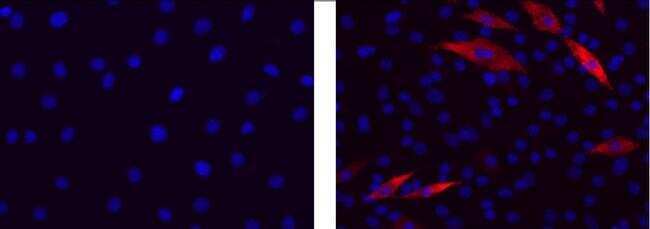
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
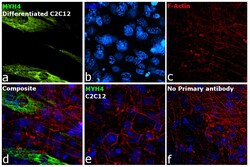
- Experimental details
- Immunofluorescence analysis of MYH4 was performed using 70% confluent log phase C2C12 cells and C2C12 differentiated Myotubes. The cells were fixed with 4% paraformaldehyde for 10 minutes, permeabilized with 0.1% Triton™ X-100 for 10 minutes, and blocked with 2% BSA for 45 minutes at room temperature. The cells were labeled with Myosin 4 Monoclonal Antibody (MF20), eBioscience™ (Product # 14-6503-82) at 1:100 dilution in 0.1% BSA, incubated at 4 degree celsius overnight and then labeled with Donkey anti-Mouse IgG (H+L) Highly Cross-Adsorbed Secondary Antibody, Alexa Fluor™ Plus 647 (Product # A32787, 1:2000 dilution), for 45 minutes at room temperature (Panel a: Green). Nuclei (Panel b: Blue) were stained with ProLong™ Diamond Antifade Mountant with DAPI (Product # P36962). F-actin (Panel c: Red) was stained withAlexa Fluor™ 488 Phalloidin (Product # A12379, 1:300).Panel d represents the merged image showing Cytoskeleton localization. Panel e represents C2C12 undifferentiated. Panel f represents control cells with no primary antibody to assess background. The images were captured at 60X magnification.

Supportive validation
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
- NULL
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
- NULL

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
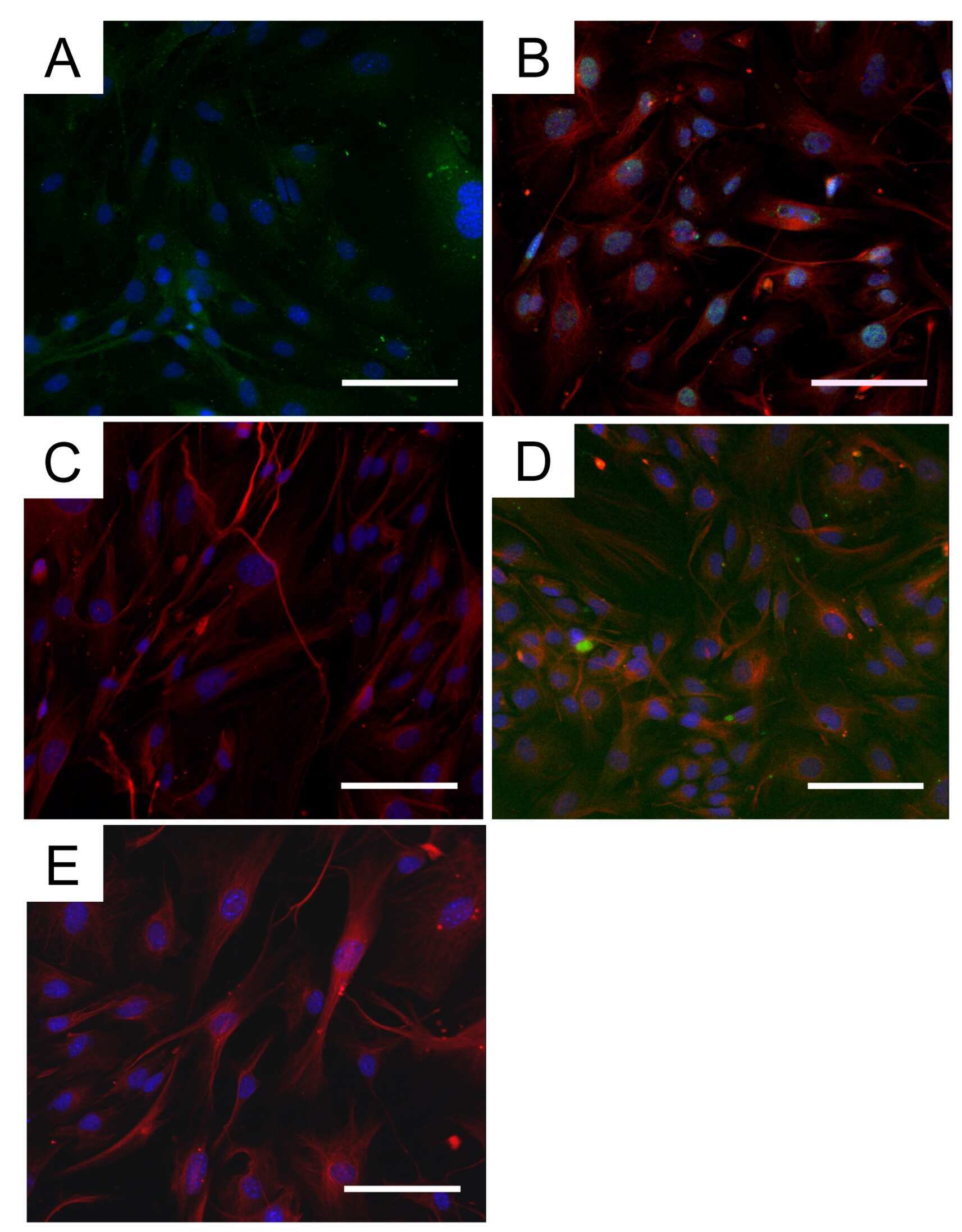
- Experimental details
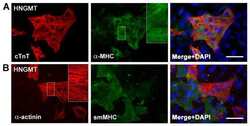
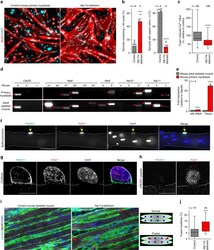
- Fig. 1 Defective myotube formation by CDC50A-deficient and ATP11A-deficient myoblasts. a - d Aberrant morphologies of PS flippase-deficient myotubes. a Syncytia formed by WT and CDC50A-deficient or ATP11A-deficient C2C12 myoblasts were visualized by immunofluorescent staining with anti-MyHC antibody (differentiated cells, red) and DAPI (nuclei, cyan). b Left: cell fusion evaluated as percentages of syncytia containing >=50 nuclei in a . Right: polarized elongation evaluated as percentages of syncytia with aspect ratios >=3 in a . c Syncytia formed by human primary myoblasts transfected with control, CDC50A or ATP11A siRNA were visualized by immunofluorescent staining with anti-MyHC antibody and DAPI. d Left: cell fusion evaluated as percentages of syncytia containing >=16 nuclei in c . Right: polarized elongation evaluated as percentages of syncytia with aspect ratios >=3 in c . e , f Rescue of morphologies in PS flippase-deficient myotubes by overexpression of PS flippase complex components. e Syncytia formed by WT, CDC50A-deficient or ATP11A-deficient C2C12 myoblasts expressing FLAG-tagged CDC50A, ATP11A, ATP11B or ATP11C were visualized by immunofluorescent staining with anti-MyHC antibody and DAPI in Supplementary Figure 3b . Left: cell fusion evaluated as percentages of syncytia containing >=50 nuclei. Right: polarized elongation evaluated as percentages of syncytia with aspect ratios >=3. f Localization of FLAG-tagged proteins expressed in CDC50A-deficient or ATP11A-def

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
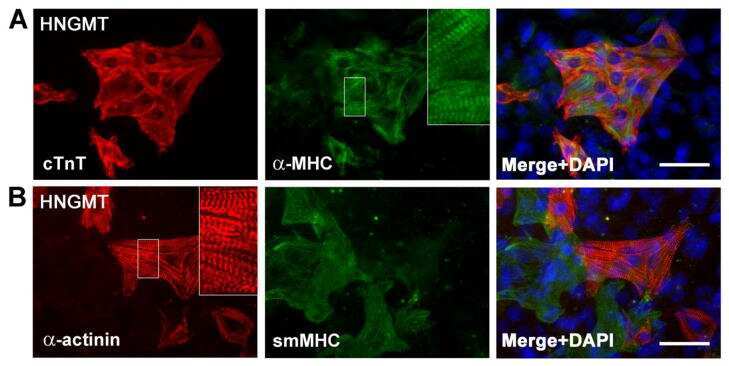
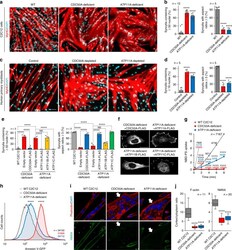
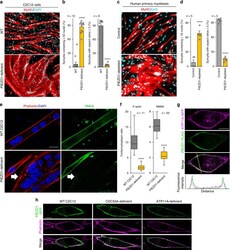
- Fig. 2 Defective myotube formation by PIEZO1-deficient myoblasts. a - d Aberrant morphologies of PIEZO1-deficient myotubes. a Syncytia formed by WT and PIEZO1-deficient C2C12 myoblasts were visualized by immunofluorescent staining with anti-MyHC antibody (differentiated cells, red) and DAPI (nuclei, cyan). b Left: cell fusion evaluated as percentages of syncytia containing >=50 nuclei in a . Right: polarized elongation evaluated as percentages of syncytia with aspect ratios >=3 in a . c Syncytia formed by human primary myoblasts transfected with control or PIEZO1 siRNA were visualized by immunofluorescent staining with anti-MyHC antibody (differentiated cells, red) and DAPI (nuclei, cyan). d Left: cell fusion evaluated as percentages of syncytia containing >=16 nuclei in c . Right: polarized elongation evaluated as percentages of syncytia with aspect ratios >=3 in c . e , f Mislocalization of cortical actomyosin in PIEZO1-deficient myotubes. e Localization of F-actin (phalloidin, red) and NMIIA (anti-NMIIA antibody, green) at the cell periphery of WT and PIEZO1-deficient C2C12 syncytia. Arrows indicate PIEZO1-deficient syncytia with diminished peripheral accumulation of NMIIA. f Cortex vs. cytoplasm ratio of F-actin and NMIIA signals in e . g , h Normal cell surface expression of PIEZO1 in PS flippase-deficient myoblasts. g Co-localization of GFP-tagged ATP11A (magenta) and FLAG-tagged PIEZO1 (green) in WT C2C12 myoblasts. Merged images and signal intensities are shown in the
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
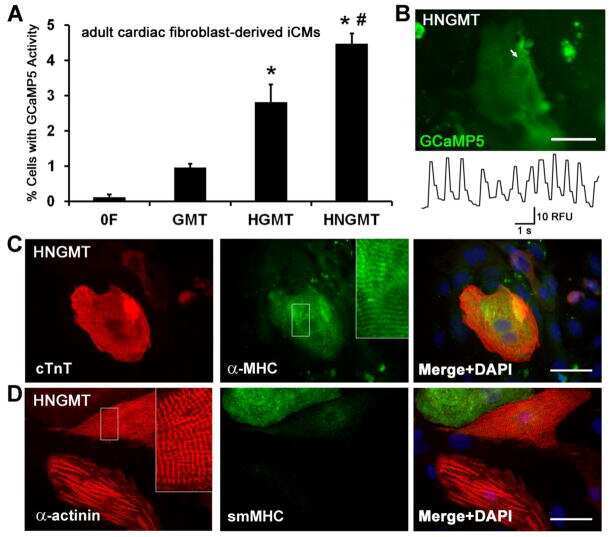
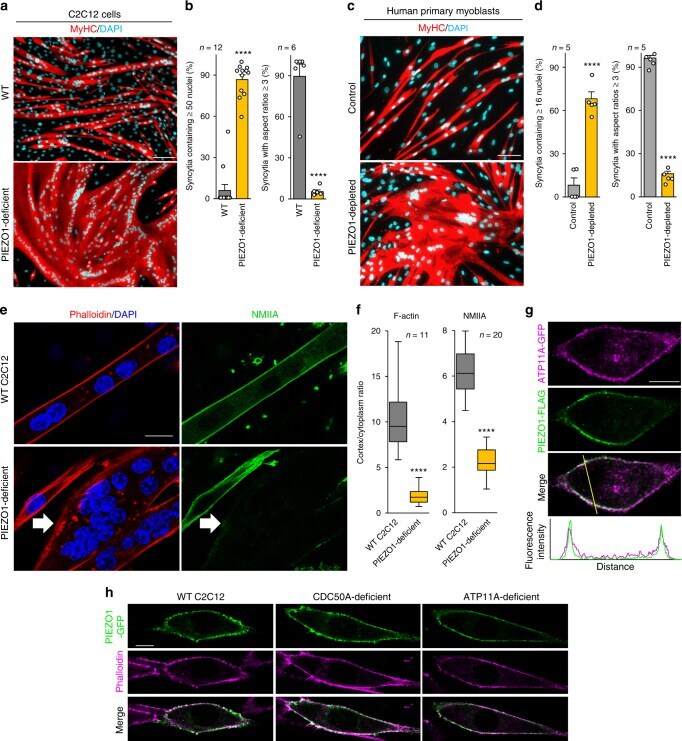
- Fig. 7 RhoA/ROCK-mediated actomyosin formation via the PS flippase/PIEZO1 pathway required for myotube morphology. a Schematic diagram of the PS flippase/PIEZO1 axis (black) and activators for the RhoA/ROCK/actomyosin pathway (red). b , c Suppressed cortical accumulation of phosphorylated MLC2 (P-MLC2) in PS flippase-deficient and PIEZO1-deficient myotubes. b Localization of P-MLC2 (anti-P-MLC2 antibody, green), F-actin (phalloidin, red), and nuclei (DAPI, cyan) in WT, PIEZO1-deficient, CDC50A-deficient, and ATP11A-deficient C2C12 syncytia. Intense co-localization of P-MLC2 and F-actin was observed on the cell periphery of WT myotubes. c Cortex vs. cytoplasm ratio of P-MLC2 signals in b . d , e Rescue of myotube formation by stable expression of a phospho-mimetic form of MLC2 in PIEZO1-deficient C2C12 syncytia. d Syncytia formed by WT and PIEZO1-deficient C2C12 myoblasts stably expressing WT MLC2 (MLC2-WT-GFP) or phospho-mimetic MLC2 (MLC2-DD-GFP) were visualized by immunofluorescent staining with anti-MyHC antibody (differentiated cells, red) and DAPI (nuclei, cyan). e Left: cell fusion evaluated as percentages of syncytia containing >=50 nuclei in d . Right: polarized elongation evaluated as percentages of syncytia with aspect ratios >=3 in d . f Rescue of myotube formation by activation of the RhoA/ROCK/actomyosin pathway in PS flippase- or PIEZO1-deficient C2C12 syncytia. Syncytia formed by WT, PIEZO1-deficient, CDC50A-deficient, and ATP11A-deficient C2C12 syncytia after

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
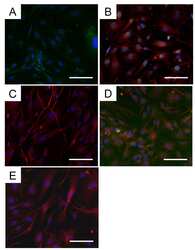
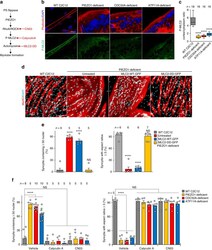
- Fig. 8 A role of ATP11A in morphogenesis during myofibre regeneration. ( a , b ) a Aberrant morphologies of Atp11a -deficient primary myotubes. Syncytia formed by control or Atp11a -deficient mouse primary myoblasts were visualized by immunofluorescent staining with anti-MyHC antibody (differentiated cells, red) and DAPI (nuclei, cyan). b Left: cell fusion evaluated as percentages of syncytia containing >=26 nuclei in a . Right: polarized elongation evaluated as percentages of syncytia with aspect ratios >=3 in a . c Suppression of agonist-induced PIEZO1 activation in Atp11a -deficient mouse primary myoblasts. Quantification of Yoda1-induced Ca 2+ influx in control or Atp11a -deficient mouse primary myoblasts. d Semi-quantitative RT-PCR analysis of CDC50 family members and P4-ATPases in mouse primary myoblasts (upper panels) and adult skeletal muscle (lower panels). Arrowheads denote specific bands. e Expression levels of Piezo1 in mouse primary myoblasts and adult skeletal muscle, evaluated by quantitative RT-PCR. Gapdh and 18S rRNA were used as internal controls. f - h Detection of endogenous PIEZO1 protein in satellite cells. Epifluorescence ( f ) and confocal ( g , h ) images of mouse myofibres visualized by immunofluorescent staining with anti-PIEZO1, anti-PAX7 antibodies and DAPI (nuclei). Specificity of the anti-PIEZO1 antibody was confirmed by the decreased fluorescence signal in the sample treated with the epitope peptide ( h ). Arrows indicate Pax7-positive satellit
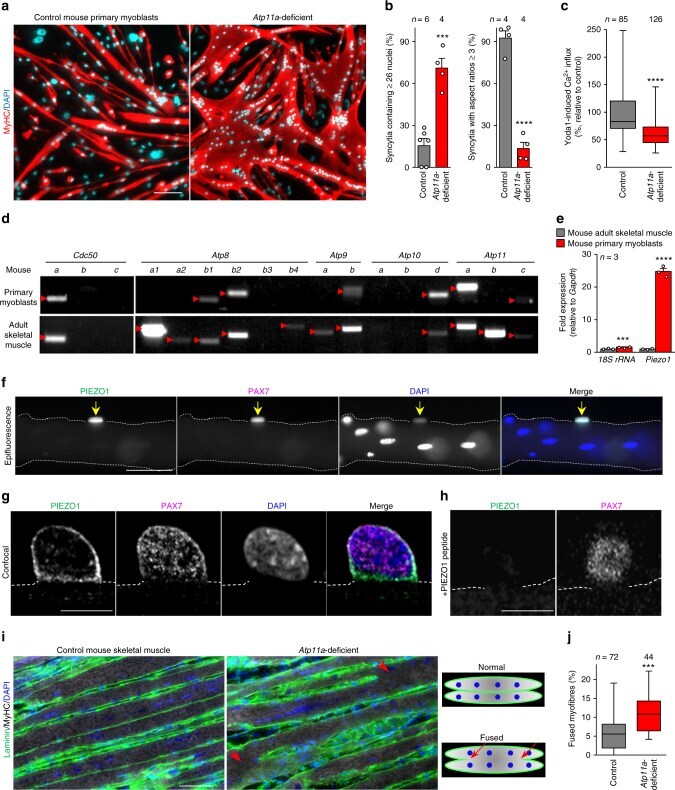
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
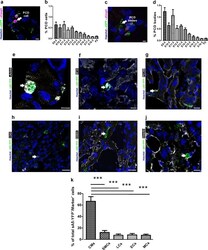
- Fig. 8 Identification and quantitation of the different cell types undergoing PCD during heart development. a Sagittal section of a transgenic heart at E12.5 depicting PCD cells. PCD cells were defined by condensed nuclei, a clear ring-like membrane sA5-YFP accumulation, and/or cCasp3 + signal. b Quantitation of PCD cells (white arrow in a). c Sagittal section of a transgenic heart at E12.5 depicting bodies of PCD cells. PCD Bodies were defined as smaller particles (2-4 um) without nuclear signal and positive for either sA5-YFP or cCasp3 or both. d Quantitation of PCD bodies (white arrow in c ). e - j Co-stainings in sections of transgenic embryonic hearts (E13.5) stained for cardiomyocyte (CMs, alpha-actinin, and MF20), endothelial cell (ECs, PECAM), leukocyte (LCs, CD45), smooth muscle cell (SMCs, ASMAC), and mesenchymal cell (MCs, vimentin) markers. k Quantitation of the different cell types that undergo PCD during heart development. Hoechst nuclear staining and sA5-YFP secreted human Annexin V-yellow fluorescent protein. Bars are 5 um ( a , c , e , f ), 10 um ( g , j ), and 20 um ( h , i ). Data are shown as mean +- s.e.m.,*** P < 0.001 ANOVA with Bonferroni post hoc test; n = 7-13

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
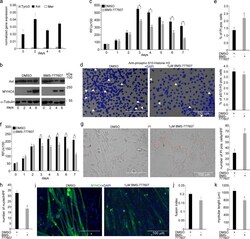
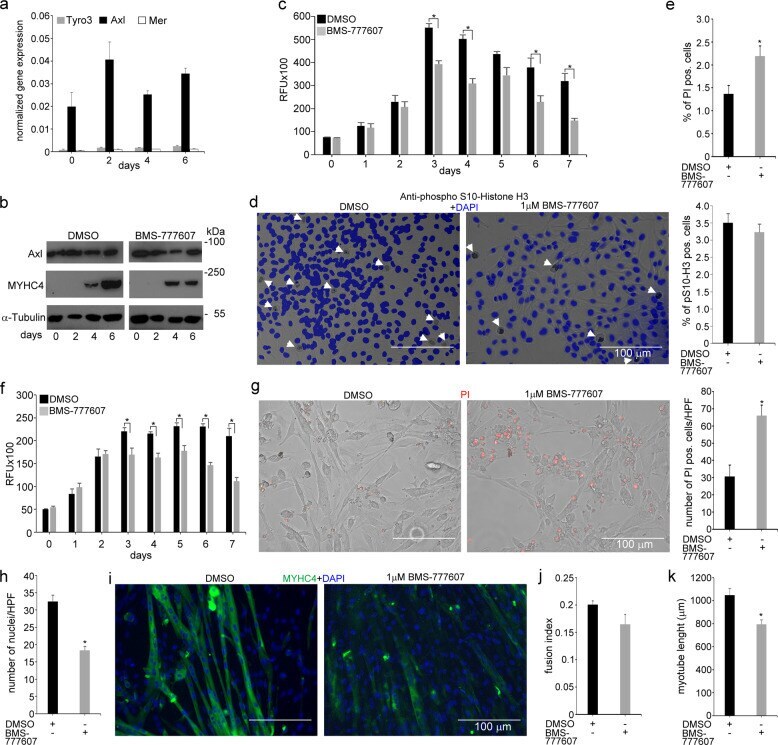
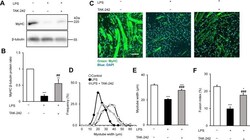
- Fig. 6 In vitro administration of the pan-TAM tyrosine kinase inhibitor BMS-777607 impairs myogenesis of C2C12 myoblast cells. a mRNA expression levels of Tyro3, Axl, and Mer during the differentiation of C2C12 myoblast cells determined by qRT-PCR. b Protein expression levels of Axl and myosin heavy chain 4 (MYHC4) in differentiating C2C12 myoblasts in the presence or absence of 1 muM BMS-777607 determined by Western blot analysis. alpha-Tubulin was used as a loading control. One representative blot of three is shown. c Alterations in the number of viable C2C12 cells grown in growth medium in the presence or absence of 1 muM BMS-777607 determined by PrestoBlue staining. d Percent of cells in G2/M phase grown in growth medium in the presence or absence of 1 muM BMS-777607 as indicated by anti-phospho-histone H3 (Ser10) and DAPI co-staining (at least 20 HPF were analyzed). Arrows point to the anti-phospho-histone H3 positive nuclei. e Percent of PI-positive cells grown in growth medium in the presence or absence of 1 muM BMS-777607. f Alterations in the number of viable C2C12 cells grown in differentiation medium in the presence or absence of 1 muM BMS-777607 determined by PrestoBlue staining. BMS-777607 was added when the medium was changed to differentiation medium. g Representative light microscopic images of C2C12 myoblasts differentiated for 6 days in the absence or presence of 1 muM BMS-777607 after staining dead cells with propidium iodide and quantification of propidium
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
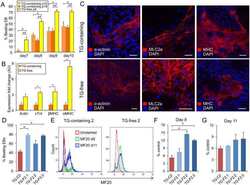
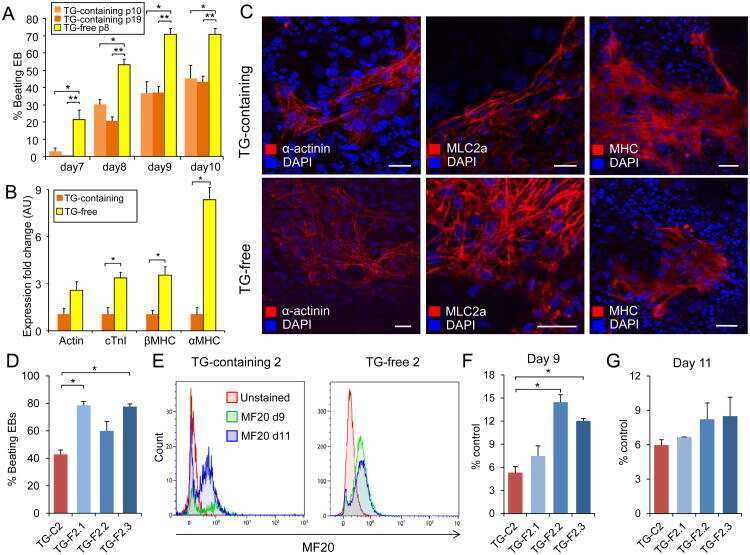
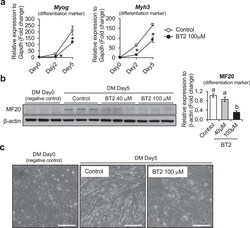
- Fig. 2 The effect of BT2 on C2C12 myoblast differentiation. a) Time-course qRT-PCR analysis of the effect of BT2 treatment (100 muM) on the expression of myogenic differentiation markers ( Myh3 and Myog ) of C2C12 myoblasts. mRNA expression levels were calculated relative to Gapdh and the data are expressed as a fold-increase. Significance was determined with the two-tailed Student's t-test (vs. control, *p < 0.05) (n = 6). Values are expressed as means +- SEM. b) The effect of BT2 treatment (40 muM and 100 muM) on total MyHC expression (anti-MF20) of C2C12 myoblasts for 5 days after induction of differentiation. Myoblasts at DM 0day (cultured in growth media) is used as negative control. Graph shows the relative intensity of each band after normalization to beta-actin. Different superscripts indicate a significant difference between 2 groups. All assessments of significance were performed with 1-way ANOVA with Tukey post hoc test (p < 0.05) (n = 3). Values are expressed as means +- SEM. c) Representative images of Control and BT2-treated (100 muM) C2C12 myoblasts for 5 days after induction of differentiation. Myoblasts at DM day0 was shown as negative control. Bar = 100 mum. Fig 2

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
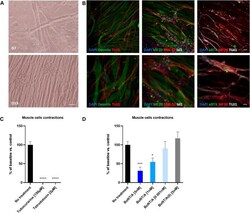
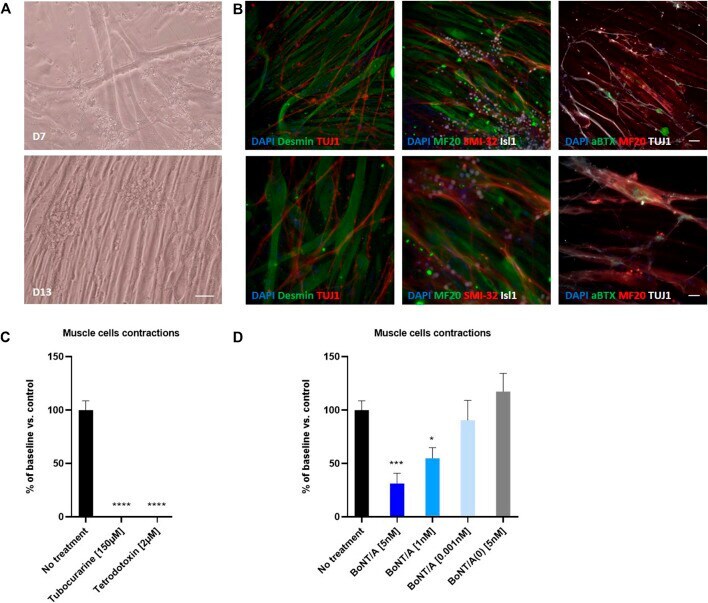
- FIGURE 7 Coculture of human myotubes and human motor neurons. (A) Phase images of co-cultures maintained in P96 during 7 or 13 days. Scale bar: 25 um. (B) Immunocytochemical characterization of NMJ formation. Muscular fibers are identified with Myosin/MF20 or Desmin, Motor Neurons neurites are identified with TUJ1 or SMI-32 and MN nuclei are identified with Islet 1 and Acetylcholine receptor with alpha-bungarotoxin. Nuclei are represented in blue (DAPI). Top: 20X images; scale bar: 50 um. Bottom: 40X images; scale bar: 25 um. (C) Effect of TTX and Tubocurarine on muscle cells contractions after 6 h of exposure. (D) Dose-response effects of BoNT/A on contraction of myotubes after 6 h of exposure. * p < 0.05; ** p > 0.01; *** p < 0.001; **** p < 0.0001.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
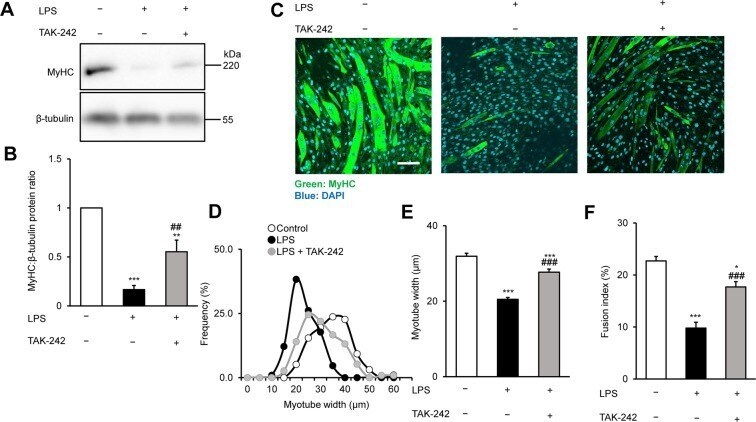
- Figure 1 TAK-242 reduces LPS-induced atrophy of C2C12 myotubes. ( A,B ) Western blot analysis ( A ) and quantification ( B ) of MyHC expression in C2C12 myotubes treated for 48 h with vehicle, LPS (1 mug/mL), or LPS and TAK-242 (1 muM). Data in ( B ) were normalised to beta-tubulin protein levels, and the ratio in vehicle-treated control cells was set to 1. N = 5/group. Full-length blots are presented in Supplementary Figure S4. ( C ) Representative immunofluorescence staining of MyHC in C2C12 myotubes treated as described for ( A,B ). Scale bar = 100 mum. ( D-F ) Distribution of myotube widths ( D ), mean myotube width ( E ), and fusion index ( F ) of C2C12 cells treated as described in ( A,B ). Myotube width was measured as described in the Methods. N = 97-114. The fusion index was calculated from five randomly selected fields as described in the Methods. For all panels, data are presented as the mean +- s.e.m. ***p < 0.001, **p < 0.01, *p < 0.05 vs vehicle control; ### p < 0.001, ## p < 0.01 vs LPS-treated group. P-values were derived from one-way ANOVA followed by Tukey's honest significant difference test or Kruskal-Wallis test followed by Dunn's post hoc tests with Bonferroni correction.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image

- Experimental details
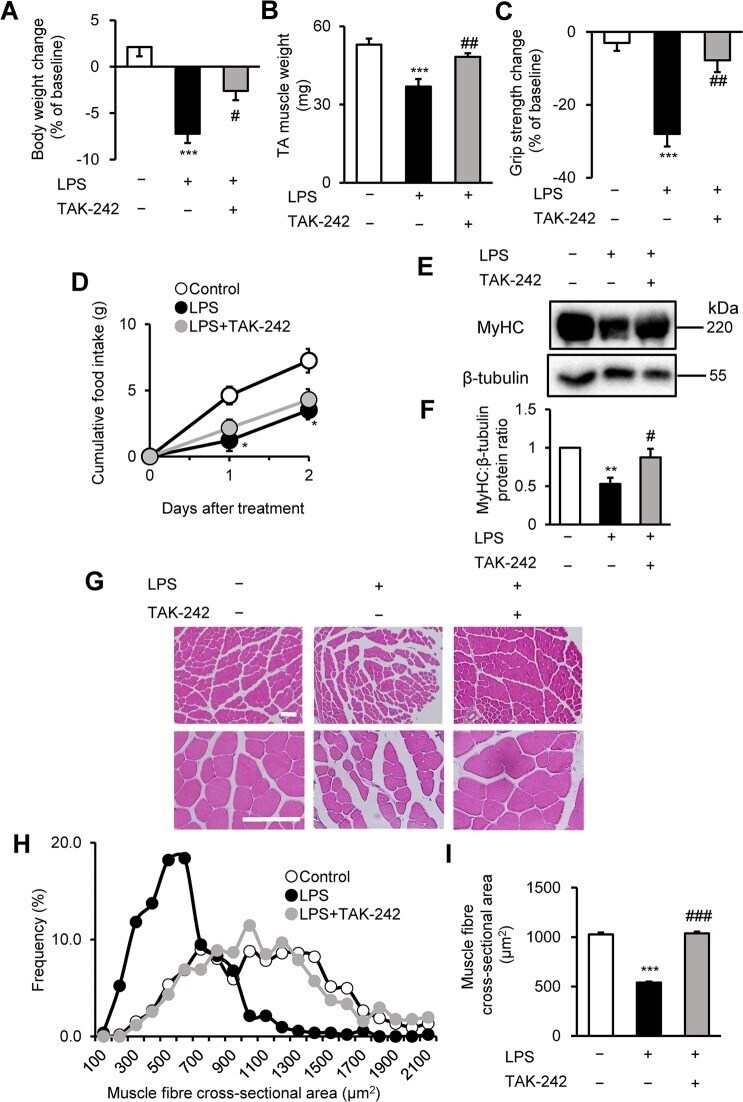
- Figure 3 TAK-242 reduces LPS-induced muscle atrophy and weakness in mice. ( A - D ) Wild-type C57BL/6 mice (8-12-week-old males) were injected with vehicle (PBS containing 0.9% DMSO) or TAK-242 (3 mg/kg) and then with PBS or LPS (1 mg/kg) 1 h later. After 2 days, mice were assessed for body weight ( A ), TA muscle weight ( B ), and grip strength ( C ). Food intake ( D ) was measured every 24 h up to 48 h. N = 4-5/group. (E , F ) Western blot analysis ( E ) and quantification ( F ) of MyHC expression in TA muscles at 2 days after administration of LPS (1 mg/kg) and TAK-242 (3 mg/kg) as described for ( A-D ). Data were normalised to beta-tubulin protein levels, and the ratio in vehicle control-treated mice was set at 1.0. N = 7-8/group. Full-length blots are presented in Supplementary Figure S6 . ( G-I ) Representative images of H&E-stained TA muscle sections ( G ) and quantification of the distribution ( H ) and mean ( I ) cross-sectional areas of TA muscle fibres at 2 days after administration of LPS (1 mg/kg) and TAK-242 (3 mg/kg) as described for ( A-D ). The cross-sectional area of TA muscle fibres was measured as described in the Methods. Scale bar, 100 muM. N = 506-523/group. For all panels, data are presented as the mean +- s.e.m. ***p < 0.001, **p < 0.01, *p < 0.05 vs vehicle control, ### p < 0.001, ## p < 0.01, # p < 0.05 vs LPS-treated group by one-way ANOVA followed by Tukey's honest significant difference test.
- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
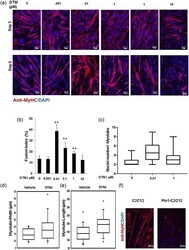
- Figure 2 Loss of peptidyl-prolyl cis-trans isomerase NIMA interacting 1 (Pin1) enhances muscle fusion in vitro. (a) Low dose of the Pin1 inhibitor dipentamethylene thiuram monosulfide (DTM; 0.01 muM) treatment caused more enhanced fusion than higher dosage (1 muM) in C2C12 cells. C2C12 cells were differentiated for 5 days with different dosage of Pin1 inhibitor DTM ranging from 0.001 to 10 muM. Cells were then immunostained for myosin heavy chain (MyHC) with anti-MyHC antibody and Cy3-conjugated secondary antimouse antibody. Scale bar= 50 muM in Day 3 images and scale bar= 100 muM in Day 5 images. (b) DTM treatment at 0.01 muM concentration shows highest fusion index. Graph showing the fusion index calculated by dividing the total number of nuclei in multinucleated cells (which have more than two nuclei) in each field by the total number of nuclei and making percentage from cells treated with DTM or without DTM ( **p < 0.001). DTM treatment at 0.01 muM concentration shows the highest fusion index. (c) Nuclei number in each myotube ( x -axis in the graph) of cells treated with different dosages of DTM. Statistics was calculated with GraphPad Prism software, and p < 0.01. (d) Myotube width and (e) length shown graphically in a box plot made with data derived from DTM (0.01 muM)-treated C2C12 cells after 3 days of differentiation. Pin1 inhibition increases both the width and length of myotubes. (f) Overexpression of Pin1 inhibiting myotube formation. Pin1-overexpressing st

- Submitted by
- Invitrogen Antibodies (provider)
- Main image
- Experimental details
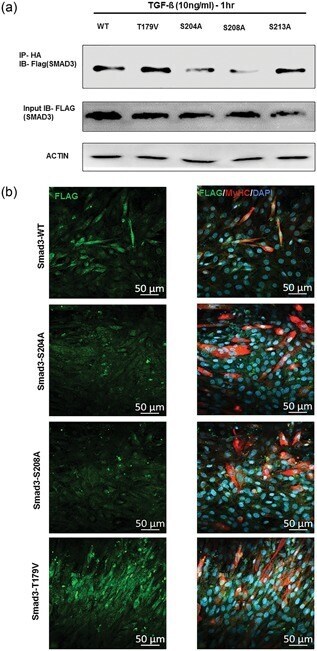
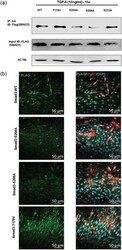
- Figure 6 Peptidyl-prolyl cis-trans isomerase NIMA interacting 1 (Pin1)-binding serine 204 residue in the smad3 linker region is crucial for fusion. (a) Pin1 binds to serine 204 and serine 208 of the Smad3 linker region in postmitotic C2C12 cells. Smad3 wild type and SP/TP site mutants in the linker region of Smad3 (CS2 Smad3-WT, CS2 Smad3 T179V, CS2 Smad3 S204A, CS2 Smad3 S208A, and CS2 Smad3 S213A) were coexpressed with PCNDA3.1 HA-Pin1. 24 hr posttransfection and postculture in a differentiation medium, cells were harvested and Immunoprecipitatioin (IP) was done with the anti-HA antibody. Later, western blot analysis was done for IP samples and input samples followed by immunoblotting for the Flag tag and actin. (b) Cells overexpressing CS2 Smad3 S204A only showed rescue of myotube fusion, and cells overexpressing CS2 Smad3 T179V showed many cells with two nuclei. C2C12 cells were transfected with constructs for Smad3 wild type and SP-TP site mutants in the linker region of Smad3 (CS2 Smad3-WT, CS2 Smad3 S204A, CS2 Smad3 S208A, and CS2 Smad3 T179V). The cells were allowed to differentiate for 4 days before fixation and immunostained with the Anti-MyHC antibody and Anti-Flag antibody. Scale bar= 50 muM [Color figure can be viewed at wileyonlinelibrary.com]